1. 引言
华南地区晚古生代发育有一套下古生界地层,为典型的浅海相沉积。研究表明,加里东期构造运动之后,华南地区泥盆纪开始重新沉降,沉积陆相至海相物质,早石炭世海侵范围略有扩大,到晚石炭世时海侵显著加大,乃至在二叠纪时期华南板块遭受了晚古生代最大规模的海侵 [1]。全球海陆分布在该时期也发生了重大的变化,在古生代末期,劳亚大陆和冈瓦纳大陆的西侧相互连接,形成了全球统一的巨型联合古陆,而在此期间华南板块始终处于海相沉积环境。
晚古生代时期古生物也发生了重要的转变。与早古生代相比,晚古生代生物发生了显著的变化。该时期陆相动植物发展迅速,但在华南地区的古生物还是以海生无脊椎动物为主。早古生代繁盛的笔石几乎完全绝灭,三叶虫也大量减少,而珊瑚、腕足、䗴类、菊石在该时期大量繁盛并占据了主要地位,在生物地层学中起显著作用。以腕足类和珊瑚类为例 [1]:
泥盆纪腕足类以石燕贝类特别发育,石炭纪和二叠纪长身贝类开始繁盛,石燕贝类和扭月贝类持续发展,其中以长身贝类最为重要。晚泥盆世的典型腕足类生物为小嘴贝类Yunnanellina。早石炭世有 Echoristites,Gigantoproductus。
晚古生代期间珊瑚类极度繁盛,尤其是四射珊瑚出现了三期发展高潮。泥盆纪为四射珊瑚发展的第一高潮期,主要代表有拖鞋珊瑚、六方珊瑚等;早石炭世是四射珊瑚的第二高峰期,出现了大量具有轴部构造的三带型珊瑚,具有重要的分带意义,主要代表有假乌拉珊瑚、泡沫柱珊瑚、袁氏珊瑚和贵州珊瑚。早二叠世珊瑚再度繁盛,为第三高峰期 [1]。
泥盆纪内部至少由8次明显的生物灭绝事件,其中弗拉斯–法门阶事件(F-F事件)较为重要,为显生宙五次生物大灭绝事件之一。石炭纪是中国地质历史上第一个重要的成煤时期,海陆变迁和陆地森林首次大规模出现,使得华北地区出现煤层,但华南地区仍为陆相沉积。二叠纪末期则发生了显生宙以来规模最大的生物大灭绝事件,海生生物出现大规模灭绝 [1]。
广西地区广泛发育晚古生代地层,例如桂中地区泥盆系一直作为我国海相泥盆系的对比标准,并用于对比研究F-F事件这一处于泥盆纪–石炭纪之交的全球性生物大灭绝事件。桂林地区也较好地保存了泥盆–石炭纪地层,是研究华南晚古生代早期古生物面貌及其古地理环境的重要窗口。其中,桂林地区下石炭统大塘阶黄金组富含珊瑚、腕足类等化石,是恢复桂林地区早石炭世生物面貌,重建生物群落特征,揭示沉积古地理的理想层位。
黄金组主要岩性以深色灰岩、硅质灰岩为主,夹泥质、砂质灰岩及硅质岩,古生物化石富集且极具代表性,代表滨岸潮坪及开阔台地的沉积环境 [2] [3]。前人对该套地层的古生物化石种类也进行了大量的归类整理,并划分出化石带。本次研究将对桂林地区下石炭统黄金组的古生物种属、化石组合等方面进行分析,初步恢复生物群落的构成及生态环境,从而进一步约束区域沉积古地理的特征。
现大部分研究 [4] 根据广西泥盆系的沉积序列及灰岩的结构,分析F-F事件的成因为弗拉斯晚期发生了海侵作用,在一段时间的深水环境后,法门早期发生了一次显著的海退,短时间内的海侵–海退作用造成了生物的大规模灭绝。也有人 [5] 研究表明,贵州地区当时发生了一次海水上涌(或变浅)事件导致海水缺氧。另有说法 [6] 认为此次事件成因是小行星撞击形成了大规模海啸造成了该全球等时性事件的发生。
“柳江运动”为朱森(1932)所命名,尔后,李捷(1934)在柳江县拉堡附近的马鞍山发现下石炭统和上泥盆统的接触关系为“角度不整合”,进而确定了柳江运动。广西石油普查大队(1959)查明,柳江运动创名地点可见泥盆系和石炭系断层接触。而广西区域地质调查队(1970)展开柳州幅(1:20万)区调工作时认为下石炭统岩关阶与上泥盆统在广西大部分地区呈整合接触,在局部地区存在沉积间断。现广西壮族自治区地质矿产局(1985)认为广西泥盆纪末部分地区上升是存在的,沿用“柳江运动”一名代表广西局部地区泥盆纪末的一次上升运动 [7]。
前人关于黄金组生物地层开展了研究 [8] [9] [10],并对地层的古生物化石进行了分类和属种鉴定并划分出了生物(组合)带 [11]。前人划分出的生物(组合)带 [11] 在桂林地区由下至上依次为:
有孔虫:Globoendothyra-Dainella组合带、Florennella-Palaeoparastaffella组合带、Eoendothyranopsis- Eoparastaffella rotunda组合带、Pseudoendothyra guangdongensis-Lituotubella glomospiroides组合带。
珊瑚:Aphrophyllum带、Donophyllum带、Thysanophyllum带。
腕足类:Levitusia humerosa组合带、Delepinea subcarinata-Megachonetes papilionacea组合带。
而另一种珊瑚带划分 [12] 则将黄金组的珊瑚带分为Donophyllum带、Thysanophyllum带。但鲜有人对其进行古生物群落分析以及复原当时的生态环境和生物群落。运用生物群落的分析和对比可以对当时的沉积环境和生态环境有更深入的认识,例如,现多运用生物群落复原的方法对塔里木盆地奥陶纪生物礁滩体的古生物群落的组成的研究 [13] 或对生物大灭绝的恢复机制进行研究 [14],从而了解群落演化以及可能存在的矿产资源如油气、煤炭等资源。
2. 区域地质背景
桂林地区地层分布广泛,其中发育较全的地层为上古生界。泥盆系分布面积最大,且地层完整,有一整套沉积序列,对泥盆系的研究有很大帮助,寒武系和石炭系分布面积次之,而二叠系及侏罗系地层出露较少,仅在东南部有少量分布。出露最老的地层为寒武系,志留系、三叠系、古近系和新近系的地层在该地区缺失。本文研究的区域出露的地层有中泥盆统、上泥盆统、下石炭统地层及第四系沉积物,其中重点研究的下石炭统分布于桂林地区的东南部(图1) [15]。
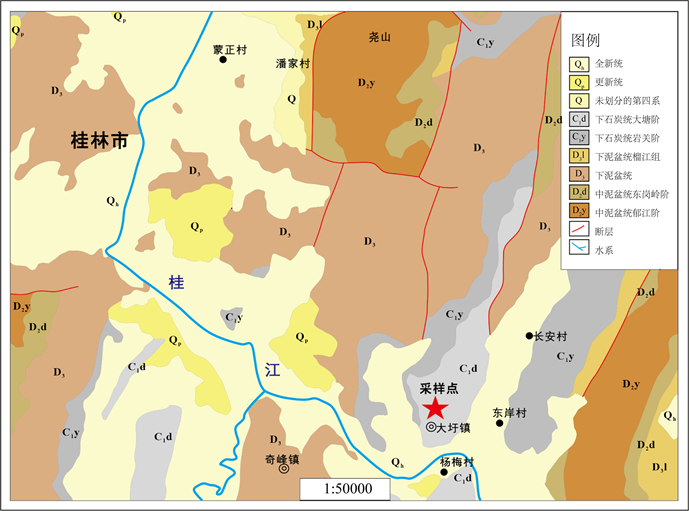
Figure 1. Regional geological map of Guilin area (according to literature [7] )
图1. 桂林地区区域地质图(据文献 [7] )
在本文研究的下石炭统中可见地层不整合现象,据前人研究 [16] [17] 在泥盆系和石炭系地层间有因“柳江运动”存在平行不整合现象,即杜内阶与法门阶的平行不整合,且石炭系各组之间也存在不整合现象。杜内阶有数个平行不整合,最明显的为尧云岭组与英沟峒组的平行不整合和英塘组与尧云岭组的平行不整合。前者分布最广,后者具有火山活动。
维宪阶与杜内阶平行不整合在广西普通可见。其位置一般在黄金组与英塘组,或都安组与隆安组之间。其上、下化石带的主要分子虽一般不跨阶,但化石带时限较长(2~6 Ma),故生物带的缺失往往不很明显,但不整合的宏观特征却较清楚。英塘组顶部常见白云岩化、角砾化、栉壳状方解石发育,顶面溶蚀构造清晰,台缘常有缺失,如桂林华侨农场鸡冠山英塘组厚234米,往东约2 km的磨盘山英塘组地层厚仅85米 [15]。
维宪阶主要有2个平行不整合,即黄金组和寺门组的平行不整合与寺门组和罗城组的平行不整合。寺门组与黄金组的平行不整合大致在珊瑚Thysanophyllum带与Donophyllum带之间,这个平行不整合导致了沉积环境的显著变化,近岸地带由碳酸盐台地沉积变为滨海含煤陆屑沉积。近岸台地由开阔台地变为浅海陆棚。这个平行不整合可称为寺门运动 [15]。
故黄金组与上覆地层(寺门组)及下伏地层(英塘组)之间均有平行不整合现象。
3. 样品特征
3.1. 样品采集
野外踏勘实测了下石炭统大塘阶黄金组的剖面:大圩钓鱼山剖面(A)、磨盘山码头剖面(B)和兴安南源村剖面(C)这三条剖面均代表了早石炭世黄金组。
大圩钓鱼山剖面(A)中可见黄金组的主要岩性为灰白色泥晶灰岩、深灰色亮晶灰岩及泥质灰岩夹少量泥灰岩,且灰岩和泥质灰岩呈韵律出现。产出大量化石,珊瑚 Syringopora,Kwangsiphyllum,Kueichouphyllum和Caninia;腕足类Gigantoproductus sp.,Productus,Martinia,Yunnanellina;和少量层孔虫及海百合茎。部分层位可见腕足碎片及珊瑚碎片,剖面底部可见异地埋藏的Kwangsiphyllum化石群及腕足碎片,剖面自上而下腕足数量和种类逐渐变多。
磨盘山码头剖面(B)中黄金组岩性主要为深灰色-灰色微晶灰岩夹泥质灰岩。产出大量化石,珊瑚Kueichouphyllum sinense,Lithostrotion (Siphonodendron) cf. petalaxoidea,Arachnolasma sp.,Clisiophyllum sp.,Dibunophyllum sp.,Thysanophyllum,Kwangsiphyllum permicum等;腕足类 Productus,Martinia,Gigantoproductus sp.等;有孔虫和海百合茎。
兴安南源村剖面(C)中黄金组岩性主要为灰黑色灰岩、灰黑色页岩、灰白色灰岩,部分含硅质、钙质结核。产珊瑚、腕足类、菊石及层孔虫等化石。
3.2. 研究区古生物特征
根据野外踏勘采集到的化石标本,可见研究区主要的化石种类为四射珊瑚、腕足类。还有少量有孔虫、海百合茎及菊石。部分化石由于石化严重且不完整,仅能鉴定到属。现结合《中南地区古生物图册》 [18] 总结描述如下:
3.2.1. 珊瑚
笛管珊瑚目SYRINGOPORIDA Sokolov,1947,
笛管珊瑚科SYRINGOPORIDA Nicholson,1879,
笛管珊瑚属Syringopora Goldfuss,1826。
丛状群体,由许多近似平行的圆柱形个体组成。个体直径1.5~2.8毫米,间距0~1毫米。个体间由连接管连接。连接管几乎均为不规则分布。横板呈漏斗状,部分具轴管。体壁薄。隔壁刺不明。
皱纹珊瑚目RUGOSA Edwards et Haime,1850,
小石柱珊瑚科LITHOSTROTIONELLIDEA Shrock et Twenhofel,1953,
泡沫柱珊瑚属Thysanophyllum Nicholson et Thomson,1876,
emend Yü,1962,
大型泡沫柱珊瑚(新种)Thysanophyllum magnum Xu(sp.nov.)。
丛状群体。成年期个体特大,个体直径15~19毫米,一级隔壁24~28个。中轴不稳定或缺失。边缘泡沫带发育。一级隔壁短,不达外壁;二级隔壁短甚至缺失。横板完全,平凸,两侧下垂。幼年未出现边缘泡沫带;青年期发育泡沫带;成年的泡沫带宽达半径的1/2。
皱纹珊瑚目RUGOSA Edwards et Haime,1850,
限珊瑚科METRIOPHYLLIDAE Hill,1939,
灰壁珊瑚属Stereolasma Simpson,1900,
湖南广西珊瑚(新种)Kwangsiphyllum hunanense Xu(sp.nov.)。
群体,丛状。个体直径4.5~5.5毫米,一级隔壁15~17个,长达半径的1/2,灰质加厚呈锥状,无鳞板。具横枝。横板完整,平凸,间距不等。
皱纹珊瑚目RUGOSA Edwards et Haime,1850,
犬齿珊瑚科CANINIIDEA Hill,1938,
犬齿珊瑚属Caninia Michelin in Gervais,1840。
锥柱状单体,直径14~26毫米,隔壁24 × 2个,主部的隔壁强烈加厚呈矛状。主内沟显著。横板通常完整平凸,不规则弯曲,微倾斜。
皱纹珊瑚目RUGOSA Edwards et Haime,1850,
古剑珊瑚科PALAEOSMILIIDEA Hill,1940,
贵州珊瑚属Kueichouphyllum Yü,1931。
大型单体,直径20~32毫米,锥柱状。一级隔壁很多,部分汇集中心;二级隔壁特别发育。横板不完整,短小,略向轴部升起。鳞板带宽,鳞板常呈同心状,列数多,半球形。主内沟显著。
3.2.2. 腕足类
长身贝亚目PRODUCTIDINA Waagen,1883,
长身贝超科PRODUCTACEA Gray,1840,
长身贝科PRODUCTIDEA Gray,1840,
长身贝属Productus Sowerby,1814。
贝体中等大小,壳长17~20毫米,壳宽15~18毫米,壳厚不明。腹壳顶区强凸,前半部凸隆较弱。背壳体腔区平,具围板,前方膝曲;体腔厚,拖曳部长。两壳壳面具清晰壳线;同心纹弱,具有明显放射纹。
长身贝亚目PRODUCTIDINA Waagen,1883,
长身贝超科PRODUCTACEA Gray,1840,
大长身贝科GIGANTOPRODUCTIDEA Muir-Wood et Cooper,1960,
大长身贝属Gigantoproductus Prentice,1950。
贝体巨大,壳长大于50毫米,壳壁厚,轮廓近圆形,铰合线等于壳。腹壳高凸;背壳深凹;两壳不膝曲;耳翼大。壳纹细密、弯曲,宽度不匀,有时集成纵脊;壳皱仅发育于铰合缘及耳翼附近;壳刺沿铰合缘排成行,少数散布于其余壳面。
石燕亚目SPIRIFERIDINA Waagen,1883,
马丁贝科MARTINIIDEA Waagen,1883,
马丁贝属Martinia McCoy,1844。
贝体大小不等,轮廓压圆形;铰合线短于壳宽主端钝圆。腹壳凸度一般大于背壳;喙突伸,弯曲;铰合面小,三角孔洞开;中槽前缘呈舌状向背方突伸。壳面饰同心纹;当外层壳面剥落厚,显露出连续的细放射纹。
小嘴贝目RHYNCHONELLIDA Kuhn,1949,
小嘴贝超科RHYNCHONELLACEA Gray,1848,
云南贝科YUNNANELLIDEA Rzhonsnitskaya,1959,
准云南贝属Yunnanellina Grabau,1931。
轮廓为不规则的三角形,不等双凸形,背壳强凸。腹喙尖而弯,中槽中隆发育,中槽前段形成长舌状延伸。壳面具多次分枝细密而平坦的壳纹,前部具独立发育而成的、长而粗强壳褶,褶顶平圆。
4. 古生物群落及岩相古地理分析
4.1. 埋藏特征及群落分析原理
野外调查发现,除了原地埋藏的化石外,还存在异地埋藏的化石,因此对剖面A和B获得的化石进行埋藏学上的分析。
4.1.1. 化石埋藏学原理
据童金南、殷鸿福(2010)在《古生物学》一书中定义 [19]:研究生物死亡后埋藏在沉积物中随同沉积物变为岩石而本身经石化作用形成化石过程的学科称为化石埋藏学。
由于各种原因而死亡的生物尸体堆积称为死亡群。死亡群可能属于同一生物群,也可能包括几个生物群死后的尸体。现代海滨介壳滩或冲剧到河口附近的生物尸体堆积就是死亡群的典型实例。死亡群经过外力作用的风化破坏、搬运过程中的破碎及溶蚀或被其他动物所吞食等,往往有一定的损失。一个死亡群还可能与其他死亡群相混合,然后被沉积物覆造形成埋藏群。经过搬运的异地埋藏群损失就会更大。随沉积物的成岩作用,埋藏群本身经石化作用形成化石群。
化石群如是由生物群死亡后埋藏在原生活位置的为原地埋藏,化石群的成员与原来生物群的成员一致,几乎全部未经移动,此化石群称原地化石群。若化石群中保存着原来生物群中的大部分成员,且保存着原地生活状态,但一小部分被搬运走了,这种原地埋藏的化石群称残留化石群。生物死亡后经过搬运,离开原地而成异地埋藏。形成的化石中大部分成员属同一生物群,并未经搬运,但混入了搬运来的生物,其中有同时期的,或有不同时沉积的,这种化石群叫混合化石群。有些化石群是生物全部搬运后再形成,它们可能来自几个同时代的生物群,此化石群称为搬运化石群。
原地埋藏的化石群对确定地层时代及恢复当时的环境非常有用。搬运化石群和混合化石群对研究古环境可提供有益的资料,如水流强度、水流方向、能量高低和沉积来源等 [19]。
4.1.2. 化石群落分析原理
据童金南、殷鸿福(2010)在《古生物学》一书中定义 [19]:化石群落(fossil community)是指生物群落死后原地埋藏被保存为化石的一部分。如果生物死亡后经过搬运形成异地埋藏,这样形成的化石群只能称为化石组合或集群。在化石群落中,若有部分生物已被搬运他处,则称为残留群落;若有其他生活环境的生物混入则称为混合集群;全部经过搬运而非原地生活的化石,构成了搬运集群。
化石群落的特征:
1) 与现生群落相比,化石群落中很少保存软躯体的生物,对微体化石通常也不够注意,未被认识的化石也常不计算在内,或由于技术问题违背修理出来。
2) 保存在同一层的化石群不一定就是原地生活群落。
3) 比较营养结构和分异度的差异,生态学研究的群落可以有生产者营养级,但在化石群落中找不到软躯体的生产者(如许多藻类),特别是在古生代的化石群落中。
4) 在化石群落中往往只有少数例子可以阐明生物之间的相互关系,如寄生现象、捕食现象、互惠共生等,因为这些现象很难保存下来 [19]。
4.2. 群落生态复原
4.2.1. 古生物化石分析
现用大圩钓鱼山剖面(A)中所采得的化石对黄金组地层从珊瑚、腕足类的含量,以及有无层孔虫、海百合茎、腕足碎片和异地埋藏现象进行简要分析(表1)。

Table 1. Summary of the fossil species content of the Daxu Diaoyu Mountain section
表1. 大圩钓鱼山剖面化石种类含量简表
根据表中在同组内的化石种类及数量分布分析可得出,在大圩钓鱼山剖面(A)中,Kwangsiphyllum (广西珊瑚)主要分布于底部1~10层,其上地层中较少甚至不可见,且在07层可见厚约10 cm的Kwangsiphyllum化石层(图2),化石产出形态为平卧,判断为异地埋藏,且产出化石的地层含泥质较多,与珊瑚自身生长环境有所差别。并在7~11层均发现有异地埋藏痕迹,如腕足化石碎片等,证明在黄金期早期发生了水体动荡。
单体形态的四射珊瑚在剖面的底部和顶部均有产出,在底部主要以Caninia (犬齿珊瑚)为主,顶部Kueichouphyllum (贵州珊瑚)数量增多且开始占据优势。
腕足类在剖面顶部才开始产出形态较完整的Gigantoproductus sp. (大长身贝),Productus (长身贝),Martinia (马丁贝),Yunnanellina (小云南贝)等,且Productus (长身贝)数量最多,占优势地位。

Figure 2. Top fossil layer (mirror to east) of layer 07 in section (A) Diaoyu Mountain, Daxu
图2. 大圩钓鱼山剖面(A) 07层顶部化石层(镜向东)

Figure 3. Ex situ burial features. (a) Brachiopod fragments; (b) Sea lily stems; (c) Flute coral, Guangxi coral and laminaria; (d) Brachiopods at the top of the rock pass order
图3. 异地埋藏特征。(a) 腕足碎片;(b) 海百合茎;(c) 笛管珊瑚、广西珊瑚及层孔虫;(d)岩关阶顶部腕足类
根据异地埋藏情况(图2和图3)分析,在黄金期早期和中期发生水体动荡,晚期水体逐渐稳定。早期生长的Kwangsiphyllum (广西珊瑚)受影响导致大量异地埋藏形成化石层,且岩层中含泥质较多,再次经历水体动荡后腕足类和珊瑚逐渐稳定,且岩性也从较多灰岩与泥质灰岩韵律产出转变为微晶灰岩。
4.2.2. 群落生态与沉积环境分析
根据《古生物学》(童金南,2010)定义 [19]:化石群落即指生物群落死后被原地埋藏的一部分。现根据剖面实测结果,对原地埋藏的化石(主要为珊瑚和腕足类)进行群落复原(图4)。
沃恩(Vaughan,1911)和韦尔斯(Wells,1957)对古生代珊瑚的生态因素研究认为,大多数古生代珊瑚的生态环境类似于现代礁相泻湖的珊瑚 [20]。古生代珊瑚适应的最大水约50 m,在水表波浪的影响下,光照强,生活于光亮带,每年水温的最低温度为16~20℃;水体富氧,振荡;底质清净,泥质堆积很少,有利于浮浪幼虫固着海底。腕足动物常与珊瑚、苔藓虫等生物共生,底栖于正常盐度的滨海–浅海地区 [20]。
由黄金期古生物群落复原图(图4)可见,黄金期早期 Syringopora (笛管珊瑚),Thysanophyllum (泡沫柱珊瑚),Kwangsiphyllum (广西珊瑚)和Caninia (犬齿珊瑚)呈生物礁生长于浅海底层,Thysanophyllum (泡沫柱珊瑚)和Kwangsiphyllum (广西珊瑚)数量居多,少量腕足呈平卧或固着于水底,中期发生水体动荡;晚期Syringopora (笛管珊瑚),Kueichouphyllum (贵州珊瑚)及少量Thysanophyllum (泡沫柱珊瑚)呈珊瑚礁生长于浅海地区,腕足类底栖于滨海–浅海地区,长身贝类以腹壳壳刺固着于底部。
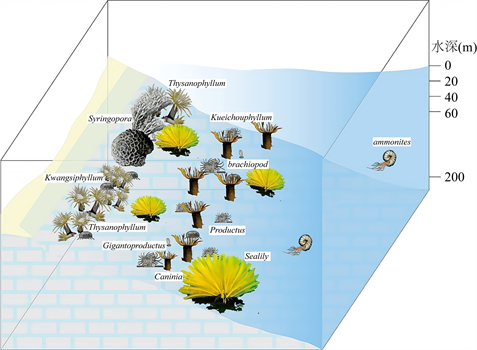
Figure 4. Recovery of the Golden Age palaeontological community
图4. 黄金期古生物群落复原图
4.3. 岩相古地理分析
研究表明,广西石炭系是在经历“柳江运动”后,地壳局部上升,海盆在相对缩小的环境中开始沉积的 [15]。但与泥盆纪具有明显的继承性关系。沉积总体反映为海进层序,在许多地方表现为由碎屑岩至泥质岩、碳酸盐岩的一个较大的沉积旋回。相序的变化也与此类似。大体上可分出滨海沼泽相、滨海陆屑滩相、局限海台地相、台沟相、台地边缘相(区)、浅海盆地相(区)等七个相,局部地段还可分出次级相。岩石组合大体上有两类:1) 浅色微–细晶白云岩、白云质灰岩夹少量灰岩、生物碎屑灰岩;2) 深色泥晶–粉晶灰岩、含燧石灰岩夹白云岩,局部夹页岩、砂岩、硅质岩等。以前一类为主。岩组厚度一般几百米,最大约千米。局部水平纹层发育。含䗴、腕足类、珊瑚等生物。
已有的研究表明,广西石炭纪古地势北东高、南西低,海盆向南西变深,海侵方向自南而北。
早石炭世岩关期,与晚泥盆世相似,桂北地区有江南古陆,东、南有鹰阳关、云开大山等陆地,这些剥蚀区是主要的物源区。当时陆源物较丰富,故滨海陆屑滩相大面积分布。与之毗邻的部分地段,形成滨海沼泽。在广泛分布的开阔海台地和局限海台地范围内,于南丹–平乐、靖西–上林、西林–百色等狭长地带出现台沟相。桂南上思、钦州、玉林一带水体较深,接受了浅海盆地相的沉积。从滨海陆屑滩相中的次级相来看,岩关期大体上经历了海进–海退–海进三个阶段,而以海退时距较长。
早石炭世大塘期,海水变浅,浅海相略有缩小,滨海沼泽相、局限海台地相和台地边缘相分布较广。从滨海沼泽相所在的地域来看,大塘期之初继续了岩关期末的海侵,陆源物少,以碳酸盐岩沉积为主;中期有较大规模的海退,海水变浅,陆源碎屑物增多,在极浅水地带形成了含煤沼泽环境;晚期海水相对加深,陆源物显著减少,碳酸盐岩沉积重新占主导地位 [9]。
5. 结论
本研究对桂林地区下石炭统黄金组三个剖面进行剖面测量、采集古生物化石并分析,现得出以下结论:
1) 下石炭统黄金组地层在桂林及周边地区岩性主要为灰岩、泥灰岩、页岩呈韵律出现。表面黄金期早期水体不稳定,较为动荡,晚期水体逐渐平静。
2) 产出的化石主要有珊瑚化石,以Kwangsiphyllum (广西珊瑚),Thysanophyllum (泡沫柱珊瑚),Caninia (犬齿珊瑚)和Kueichouphyllum (贵州珊瑚)为主;其次为腕足类化石,以Productus (长身贝)和 Gigantoproductus (大长身贝)为主,与前人研究中对早石炭世四射珊瑚的第二次繁盛期和腕足类发展特征相对应。
3) 桂林地区早石炭世黄金期的环境特征和生物演变过程如下:黄金期与岩关期之交时期Syringopora (笛管珊瑚),Thysanophyllum (泡沫柱珊瑚),Kwangsiphyllum (广西珊瑚)和Caninia (犬齿珊瑚)居多,腕足类较少,且Thysanophyllum (泡沫柱珊瑚)和Kwangsiphyllum (广西珊瑚)占优势,后在黄金期早期水体较为动荡,不利于群体珊瑚礁生长。直到黄金期晚期水体逐渐平静,腕足类开始繁盛,Gigantoproductus sp. (大长身贝),Productus (长身贝),Martinia (马丁贝),Yunnanellina (小云南贝)等均有出现,且Productus (长身贝)为优势物种。Kueichouphyllum (贵州珊瑚)在黄金期晚期逐渐替代了Caninia (犬齿珊瑚),数量增多,Thysanophyllum (泡沫柱珊瑚)数量减少,Syringopora (笛管珊瑚)变化不大。
参考文献