1. 引言
涡虫属于扁形动物门(Platyhelminthes),涡虫纲(Turbellaria),三肠目(Tricladida)。分布广泛,为淡水种,有的生活在海水、陆地或寄生。涡虫是广泛生活在洁净水质的池塘和溪流中的生物。在动物系统演化的历史中,涡虫纲占有十分重要的地位,从扁形动物开始出现了中胚层和两侧对称体制。
涡虫背腹扁平,虫体细长且柔软,体长一般0.5~2 cm左右,背部颜色深,呈黑褐色,腹面颜色浅,头呈三角形,两侧有2个明显的耳突。解剖结构复杂,包括大脑、眼睛、肌肉、肠道、神经、表皮、生殖系统 [1] 等。大脑由不同类型的神经元和胶质细胞组成,并由两条腹神经索相连。体壁肌肉系统包括纵向纤维、环形纤维和斜线纤维。表皮产生粘液,腹侧纤毛浓密,便于运动 [1]。原肾管广泛分布,用于排泄废物和调节渗透压。口位于腹面近体后的1/3处,内有肌肉质咽,是三角涡虫摄食的器官,生殖孔位于口后。围绕内脏的是间充质组织室(实质),包括成体中唯一具有分化增殖能力的细胞——neoblast [2]。雌雄同体,异体受精,体内具有雌雄生殖器官,生殖方式包括无性繁殖和有性繁殖。无性繁殖通常以横分裂的方式进行,其分裂面常发生在咽后方,有性生殖主要在夏季、冬季和初春进行。
涡虫的再生能力极强。能够再生出身体任意损伤的部位,这种功能依赖于一种称为neoblast的干细胞,是涡虫体内唯一能够分化的细胞。实质组织是分化新细胞和再生组织的主要来源。将虫体横切为2段或多段,每段均可再生出完整的涡虫。如将虫体切为3段,前段再生出尾,后段生头,中段前部再生头,后部再生尾。不仅如此,涡虫的再生过程也非常迅速,它能在一周内重新长出被切割掉的肌肉、皮肤、肠道、生殖系统,甚至整个大脑。涡虫独特的结构和极强的再生能力是研究再生、胚胎发育和细胞分化重要的实验材料,也是研究后生动物起源进化的极佳模式生物。
当动物受到损伤后会产生创伤反应,包括细胞募集、细胞增殖、免疫反应等。涡虫能够识别创伤反应和再生修复,可以完全再生出身体缺失的部分。再生需要干细胞,涡虫中的neoblast是唯一能够进行增殖分化的干细胞,由多能干细胞、γ-neoblast、σ-neoblast及不同的祖细胞亚群组成,分布全身,在截肢后,伤口处出现芽基,neoblast在创面增殖分化 [3]。这些祖细胞是neoblast标记的Smedwi-1和特定转录因子的表达来定义的,将其沉默会损害不同类型细胞的再生 [4]。
再生的细胞类型有两种:再生的潜能以及指导再生的信息。再生的潜能性指特异性成体细胞替代缺失细胞。再生的信息指的是指引哪种类型细胞再生的分子机制。位置调控信息(position control genes, PCGs)被认为是一个关键的问题。
涡虫中,多能干细胞有产生所有新组织的能力。PCGs可以调控再生以及组织的替换。皮下细胞与许多PCGs共表达(见图1)。推测皮下细胞含有机体平衡和再生的位置调控。所有涡虫的肌肉均表达PCGs,意味着涡虫的肌肉组织有两种功能:调控涡虫机体模式以及机体的收缩。受损后,PCGs在肌肉中的表达是动态的,说明肌肉对再生是有指导意义的。涡虫的再生包括两个高度灵活的系统:具有多能性的neoblast可以再生出任何新的细胞类型,肌肉可以提供位置信息 [5] [6]。
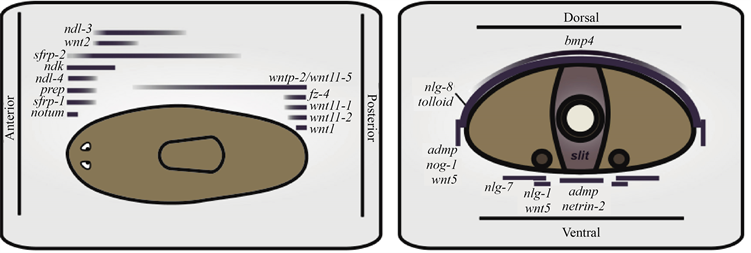
Figure 1. Cartoon depicting domains of PCG expression, which define positions across AP, DV, and ML body axes [7]
图1. 涡虫中位置调控基因在背腹轴、左右轴和前后轴中的分布 [7]
2. 涡虫肌肉系统结构
涡虫缺乏循环系统、骨骼系统和呼吸系统,被不同方向排列的表皮下肌肉纤维组成的致密而紧凑的网络所包围。腹部表皮下分布纤毛,运动基本是通过纤毛的滑行进行,肌肉可以支持这种运动,主要是用来确定运动的方向。
一些脊椎动物与无脊椎动物可以通过肌肉的形态特征区分为横纹肌或平滑肌。但涡虫没有明显的肌肉类型 [8],具有介于横纹肌和平滑肌之间的特征 [9],即有由肌丝组成的螺旋条纹,但含有致密的平滑肌小体。其结构与平滑肌相似,没有条纹但有致密的小体,且细胞核位于细胞的外围区域,有与肌膜直接相连的Z形线状结构,这是横纹肌的特征 [10]。涡虫的体壁肌肉组织,在整个机体内形成复杂的肌肉网络,分为环肌、纵肌以及斜肌。这种肌肉网络在某种程度上弥补了这些动物缺乏真正的骨骼系统,肌肉网络对由肠道、实质组织和其他器官的液体组成的静水骨骼起作用 [11]。
肌球蛋白在所有真核细胞中都高度保守,在真核细胞中为各种运动提供必要的动力,包括胞质分裂、吞噬、细胞器运动和肌肉收缩等 [12]。在涡虫中鉴定出两种不同的肌球蛋白重链(MHC)基因,编码两种不同的肌肉纤维类型。一种表达于咽部的肌纤维,胃皮周围的肌肉,遍布体壁的几个散在细胞,以及咽底部间充质的一些肌纤维 [13]。另一个MHC基因在表皮下体壁肌肉组织和背腹纤维中表达。MHC蛋白具有ATP酶活性,提供肌肉收缩所需的能量。由于肌肉纤维的收缩速度和ATPase活性随其MHC亚型组成的不同而不同,有可能涡虫MHC基因的不同表达模式赋予了涡虫肌肉纤维不同的生理特性。每种MHC亚型可能介导不同的生物学功能 [14]。
在日本三角涡虫中分离得到两个MHC基因DjMHC-A和DjMHC-B,二者都是横纹肌型,尽管在日本三角涡虫的肌肉中没有观察到横纹肌 [8]。DjMHC-A在咽部、肠管周围、咽腔和部分体壁肌肉中表达,DjMHC-B在体壁肌、垂直肌和水平方向的肌肉总表达。由此可见,DjMHC-A标记的肌肉类型可能与涡虫的蠕动运动相关,DjMHC-B标记的肌肉类型可能与身体运动相关 [8]。
3. 涡虫的极性
动物有不同的解剖结构并且大小有很大的差异,但他们有共同的信号通路,比如BMP信号,能够调节两侧对称动物的背腹(D/V)轴 [15] [16] [17] [18]。在涡虫中,切除DV轴后,BMP基因家族蛋白BMP4和ADMP可以促进组织的修复。ADMP的同源基因Smed-Admp在腹侧表达,Smed-Bmp4在背部表达。Smed-Admp可以促进Smed-bmp4的表达,Smed-bmp4抑制Smed-Admp的表达。Admp(RNAi)后,截取涡虫的头部与尾部,发现在涡虫再生的过程中出现头部与尾部的缺陷,这是BMP信号通路典型的功能障碍表型。ADMP是涡虫截断后外侧边缘、背部及腹部的组织再生所必需的。BMP信号对矢状截取的涡虫再生至关重要 [19] [20]。矢状面截肢面会出现两个左右不对称的片段:一个薄片段必须重新生成一个大小适中的BMP4表达结构域,另一个厚片段重新定位并调节BMP4表达结构域,以适应新的动物尺寸。ADMP(RNAi)能够在已存在的组织内沿前后轴再生一些结构但不能再生新的侧向标记的表达。未截取的ADMP(RNAi)表现出异常的身体尺寸。表明ADMP在动物动态平衡期间维持体型和正确的ML模式至关重要。BMP4能够促进背部组织的维持和再生。在左右不对称片段的再生过程中,ADMP促进了BMP4的表达并调控了BMP4的表达定位。
表明Bmp/Admp调节两侧对称动物的中心特征,广泛用于D/V轴的建立、维持和再生。前端伤口应答(截肢后3~18小时),创面肌细胞中的信号机制决定是否再生出一个新的头部。创伤反应迅速。Wnt信号被前端伤口选择性抑制,包括notum的激活。头部再生的第一阶段导致wnt信号在创面前端抑制,在创面后端激活。在创伤诱导的wnt1和notum表达的最初阶段,FoxD在伤口的中线表达 [21]。
Wnt/β-catenin和Hedgehog (HH)信号通路在再生和细胞的动态平衡中的前后轴(A/P)的确定中起重要的作用。位于涡虫前端和后端两个不同的区域,称为前极和后极,前极表达Notum,后极表达wnt。前端受损后,创伤反应在创伤处触发wnt的表达,wnt信号在前端选择性抑制 [22],同时Notum被激活,FoxD在多数伤口的中线激活。FoxD突出了中线与极点再生之间的联系。在中线有一小群表达Notum、Flolistin和FoxD的细胞从neoblast中出现,在再生过程中可以分化形成眼睛等 [19] [20] [21] [23]。因此创面中线附近的neoblast对基因的诱导是形成新的前极细胞的最初步骤。随着neoblast的增殖分化,头部前极模式建立,FoxD(RNAi)抑制前极的再生,虽然前极仍然有脑细胞的存在,但缺乏野生型头部芽基中正常组织结构,同时,中线表达基因的表达域也出现了异常。再生的前极促进了中线的再生,从而促进了头部的双侧模式的建立,并促进了头部A/P模式的基因结构域的建立 [22] [24] [25] [26]。
4. 涡虫的肌肉再生研究进展
在再生过程中,肌肉纤维通过收缩来闭合伤口,从而发挥早期功能。除体壁肌外,涡虫在消化系统、生殖器官,咽部周围均有肌肉纤维。涡虫的体壁肌肉系统由位于不同方向的不同数量的肌肉纤维层组成,其结构随身体大小而变化。大量的背腹纤维连接背侧和腹侧的身体表面。这些纤维在动物的尖端和边缘比在身体的中心区域更丰富。所有的纤维排列成致密、紧凑的肌网。动物前端部的背侧和腹侧的内纵纤维形态不同,背侧的纤维向前端部的中心区汇聚,腹侧纤维在接近前端时平行分布,甚至呈扇形发散的形式 [27]。
再生过程中,涡虫干细胞如何再生出肌肉系统,新的肌肉细胞如何整合到先前存在的肌肉网络中,以及控制肌源性基因程序的分子途径的识别,是研究涡虫肌肉再生的关键因素。目前需要解决肌肉再生的问题:在再生的过程中,新的肌肉如何整合到先前存在的肌肉网络中,控制成肌基因程序的分子途径的识别 [5]。
随着再生的进行,肌细胞可以沿着两条主要途径:1) 进入芽基形成新的肌纤维,2) 在芽基后区域完成分化并插入肌纤维。两种机制同时间进行,若肌细胞进入芽基,意味着芽基不仅通过neoblast的积累而生长,而且还通过不同分化阶段的neoblast的亚基向不同类型的细胞生长,这是恢复缺失组织所必需的 [27]。
涡虫中不同的肌肉纤维在再生过程中具有不同的调节作用见图2。MyoD是进化上保守的bHLH转录因子家族,在骨骼肌源性谱系的分化中发挥关键作用。研究发现,保守的转录因子MyoD在可能的肌源性祖细胞中表达,分化后的肌肉细胞存在位置调控基因,可以为涡虫的neoblast的增殖分化提供位置信息,使其在缺失的位置再生出正确的组织。MyoD指定了体壁肌细胞中的一个子集:纵向纤维,Nkx1-1编码了涡虫中的环状纤维。将MyoD(RNAi)后,抑制再生,将Nkx1-1(RNAi)后,中线分叉,再生出两个头 [28]。
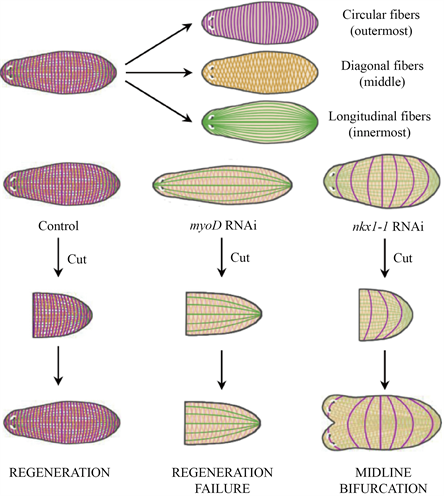
Figure 2. Planarian body wall muscle fibers and their critical roles in regeneration and patterning [28]
图2. 涡虫体壁肌纤维及其在再生中的关键作用 [28]
在涡虫中,不同的转录因子指定了不同的肌肉亚群。对揭示纵向和环状纤维的不同作用提供了重要依据。环状纤维是正常的内侧再生所必需的,在环状纤维减少后,出现两个头而不是一个头。另外,需要纵向纤维启动再生程序,缺乏纵向纤维的涡虫不能够重建肌肉中的位置信息,即使涡虫仍然具有组织更新的能力,但缺乏干细胞对损伤的反应。两种纤维类别的同时丢失则导致涡虫结构的紊乱。肌肉在涡虫的生理作用中,除了收缩,不同的肌肉纤维在协调涡虫的再生过程中起着重要的调节作用 [28]。
5. 肌肉与极性
涡虫被切割成微小片段也可以再生出完整的个体,这个过程依赖于干细胞的增殖与分化,另外也需要PCGs调控机体的构造。在涡虫的再生过程中,众多PCGs在多个体壁肌中表达。但是不同的肌肉纤维以及他们对于再生应答的作用尚不清楚。Neoblast是涡虫中未分化的细胞,具有分化出各个组织的能力。对于neoblast的增殖,涡虫的再生依赖PCGs确定再生过程中的组织特异性。受伤后,PCG在体壁肌中表达。MyoD编码肌肉发生中具有进化保守作用的螺旋–环–螺旋转录因子,MyoD(RNAi)后导致纵向肌肉纤维丢失而不干扰环形或斜线纤维,以增加涡虫的长宽比例。对未受损MyoD(RNAi)的涡虫进行RNA测序(RNA-seq)发现,PCGs的一个子集与MyoD共表达,并显著下调,说明在纵向肌肉纤维中显著表达。另外,在MyoD(RNAi)处理后的动物片段受损后不能启动再生即使neoblast有分化成各种细胞类型的能力。虽然MyoD(RNAi)可以进行小的创伤比如眼睛的修复并且通过neoblast分化进行组织的更新,但是祖细胞停止增殖以限制大的组织修复。MyoD(RNAi)RNAs-seq的动物揭示了两种伤口诱导基因的表达明显下调。两种基因通过在损伤后的纵向纤维中富集:notum和fst [29]。这两种基因在再生应答中有重要的作用:notum编码WNT脱氨酶并通过负调控wnt信号通路调节受损后涡虫头尾的结构。Fst编码follistatin并能够维持伤口诱导基因的持续表达,在再生过程中促进neoblast的增殖。当fstRNAi与MyoDRNAi相似,允许组织更替,但完全阻止再生。Myod(RNAi)与fst(RNAi)的尾部片段不能限制后部PCGs的表达,也不能正确的启动前部PCGs的表达。纵肌不仅仅是创伤后维持收缩能力所必需的,而且也是再生反应的关键调节因素 [26]。
6. 总结
涡虫的再生能力极强,即使被切割成279份,每一碎片均能再生出完整的个体。这一过程依赖于干细胞的增值与分化。干细胞是一种能够自我更新、持续分化的一类细胞,能够在动物体内产生所有类型的细胞。在涡虫中唯一具有干细胞多能性的一类细胞称为neoblast,具有异质性。
虫体受创后,再生早期会触发neoblast有丝分裂两个高峰:1) 对任何损伤的全身反应,当损伤导致组织缺失时,诱导第二个neoblast反应;2) 在伤口处招募neoblast以增殖分化出所缺失的组织。Neoblast创伤反应存在两个功能不同的信号阶段,并以此可区分受伤和组织缺失再生的情况。
创伤反应由创伤处任何细胞类型中激活的基因和体内广泛分布的细胞中激活的特定细胞类型的基因组成,共三个组成部分:1) 创伤后几乎所有类型的细胞都有基因表达,包括多个应激反应基因,2) 71%的细胞特异性基因优先在三种细胞类型(neoblast,肌肉,表皮)中的一种,转录因子和与增殖相关的基因中表达,3) 部分创伤诱导基因在肠道、咽旁细胞和神经元中表达,反映了创伤后组织独特的生理反应。在干细胞与分化组织之间,整个虫体发生转录变化。并可以通过早期反应基因分析出不同类别的创伤反应基因。预先存在在原组织中的这些早期转录因子对于启动再生是必不可少的。
受损后,涡虫会迅速愈合伤口并重新建立正确的轴向极性,在几乎任何类型的损伤中,都能够正确地再生出所缺失的部位,确定涡虫虫体适当的比例。
在涡虫发育的动态平衡中,许多模式基因在不同的结构域中表达。去头去尾都会扰乱这种正确的模式,导致不成比例的表达模式。截肢后,不同结构域的标志物会前向后向的聚集,即使在没有干细胞存在的情况下也是如此,表明这些标志物可以提供再生过程中的轴向信息。而涡虫中提供位置信息的基因在皮下肌肉细胞亚群中表达,为再生创造出一个坐标系统,通过转录激活模式基因提供位置信息。如此,neoblast可以在受损位置分化出缺失的组织与器官。
然而,肌肉是如何被指定的,不同的肌肉亚群是如何影响再生的还不得而知。
MyoD是helix-loop-helix转录因子,在肌肉发育过程中具有保守作用,与Myf5和Mrf4一起作用于脊椎动物骨骼肌的形成 [29]。在线虫中,myoD同源物hlh-1与UNC-120和HND-1协同作用,协调体壁肌肉的发育。果蝇则需要myoD同系物NAU来分化有限的肌肉 [29]。涡虫中myoD主要在体壁肌肉细胞中表达,一小部分myoD+细胞共表达neoblast标记的smedwi-1。在地中海涡虫中,只有46%的体壁肌细胞表达myoD,说明myoD可能在特定肌肉细胞亚群或短暂分化阶段表达。在涡虫中存在于沿前后轴定向的纵向纤维中。将MyoD(RNAi)后,纵向纤维缺失,环状纤维和斜线纤维保持正常,而纵向纤维丢失导致再生失败,环状纤维的形成需要不同的转录因子编码基因nkx1-1,环状纤维的缺失导致前后轴分叉。在未受损的涡虫中,myoD(RNAi)后显示,肌肉特异性基因表达减少,其他编码的保守转录因子的基因(snail,lbx,nkx6-2和lhx2/9)表达降低。这些基因在体壁肌中表达,部分与myoD一起表达,snail,lbx和nkx6-2的RNAi不会引起主要肌肉表型的变化但可以调节纵向纤维。一些与myoD共表达的PCGs基因的表达也明显降低 [29]。
myoD(RNAi)后,肌肉创伤诱导的notum和fst的表达显著减少,同时纵向纤维缺失。同时,fst和notum在myoD+细胞中的表达丰富,说明notum和fst在创伤诱导基因中的表达是独一无二的且他们的表达仅限于纵向纤维。肌肉与极性密切相关,但两者之间如何相互影响还不得而知。
NOTES
*通讯作者。