1. 前言
石漠化是由于对土地的不适当利用,使岩石裸露增加,导致土壤质量退化,生态遭到严重破坏,土地生产力严重下降的土地退化过程 [1]。我国贵州和广西等喀斯特地区水土流失、岩石暴露等土地退化问题较严重 [2]。截至2016年底,广西石漠化或潜在石漠化土地面积共为420.3万hm2,石漠化程度仅次于贵州省,与2011年相比,广西石漠化面积净减少39.3万hm2,治理已经初显成效,但与此同时,广西潜在石漠化面积反增37.6万hm2,说明导致广西土壤石漠化的因素依然存在,广西石漠化防范治理的任务依然十分艰巨 [3]。河池地处桂西北,土地面积3.35万km2,是一个水热条件良好,岩石成土速率慢,且土壤侵蚀作用强烈的溶岩地区,溶岩面积2.211万km2。同时,河池市是我国少数民族和贫困人口集中分布地区 [4]。土壤微生物在调节土壤养分、维持土壤生态系统平衡等方面具有重要的作用 [5]。微生物通过光合、化能、分解等作用影响着植物的生长,土壤中细菌和放线菌数量的增加对植物的生长发育有益,而土壤真菌化会导致土壤肥力下降,增加土壤致病力 [6],植物通过根系或凋落物向土壤环境释放各种有机物质(糖类、氨基酸类、生长因子等)和无机离子,改变土壤微生物生长的微生境,进而影响土壤微生物的群落结构。相关研究结果表明:土壤微生物种群数量、群落结构以及空间分布等,直接反映微生物的生境,同时可以客观反映土壤质量与土壤健康状况 [5] [6] [7] [8] [9]。
桂西北岩溶地区石漠化面积广、程度深,生态环境脆弱。目前,针对于桂西北石漠化的治理,学者们开展了不少研究工作,提出了一些有价值的建议,但以往的研究大部分是基于传统的手段和方法 [10] [11] [12] [13] [14]。本研究旨在通过现代技术与传统方法的结合,即将分子生物学技术与传统的土壤理化分析方法结合,比较不同植被土壤微生物多样性和主要菌群的差异,及其与土壤肥力的相关性,从而揭示桂西北岩溶地区乡土树种与外来树种对土壤质量变化及土壤微生物多样性的影响,为岩溶地区植被恢复和石漠化地区生态恢复重建提供理论参考。
2. 材料与方法
2.1. 研究区域概况
本研究的地点选择在河池市宜州区及其周边区域,地理坐标为106˚34'~109˚09'E,23˚41'~25˚37'N,属于亚热带季风气候区,为典型的溶岩地区,热量丰富、光照充足,年日照时数为1447~1600 h,气候温和,雨量充沛,年平均降雨量在1200~1600 mm,降雨主要集中在5~8月,年均气温为20.6℃,无霜期327 d。
2.2. 样品的采集及处理
2.2.1. 样地的选择
2019年5月至7月,通过实地调查,在立地条件尽量一致的情况下,分别在罗城县四把镇龙马村向阳屯,环江县明伦镇吉祥村,宜州区怀远镇叶茂村、刘三姐镇下枧村和小龙村大长沙屯、龙降村明灯寨等地,选择优势种分别为枫香、柏木、任豆、青檀、桉树、湿地松的不同植被典型样地,以上样地的编号分别为:XYFS、JXBM、YMRD、XJQT、DCSAS、MDZSS,各调查样地的基本情况见表1。

Table 1. List of different vegetation survey sample plots
表1. 不同植被调查样地情况一览表
2.2.2. 土壤样品的采集及处理
在每个调查样地,根据具体的地形地貌和植被的分布情况,用梅花形采样法采集优势树种根部周围0~20 cm深的表土。采样时,先清除采样点表面的凋落物、砾石及杂物,然后取0~20 cm的表土,过10目(孔径2 mm)标准筛,将5个小样点的土壤样品充分混合均匀,用“四分法”取样。每个样地采集的土壤样品分为两份,其中一份带回实验室自然风干后,用微型土壤粉碎机将其粉碎过筛,装入自封袋中保存,用于理化性质的测定;另一份采集后迅速放入冰盒中低温保存,最大限度保持土壤微生物原有的种群结构,带回实验室后置于−80℃的超低温冰箱中保存,用于土壤微生物DNA序列的测定。
2.3. 测定指标及方法
2.3.1. 土壤理化分析
土壤pH的测定用电位法(水土比为2.5:1) [15];全磷含量的测定用氢氧化钠碱熔——钼蓝比色法 [16];有效磷含量的测定用碳酸氢钠浸提——钼锑抗比色法 [17];全氮含量的测定用凯式定氮法 [18];速效氮含量的测定用碱解扩散法 [19];全钾含量的测定用氢氧化钠碱熔——火焰光度计法 [20];速效钾含量的测定用乙酸铵浸取——火焰光度计法 [21];有机质含量的测定用重铬酸钾氧化——分光光度法(HJ615-2011) [22]。
2.3.2. 土壤微生物DNA序列的测定
土壤微生物测序由深圳华大基因股份有限公司进行,测序区域为16S-V4,注解数据库为GreenGene数据库。首先取质量合格的基因组DNA样品及引物配置成PCR反应体系,设置PCR反应参数进行扩增,PCR扩增产物纯化后溶于Elution Buffer,贴上标签,完成建库后使用Agilent 2100 Bioanalyzer对文库的片段范围及浓度进行检测,检测合格的文库根据插入片段大小,选择HiSeq平台进行测序 [23]。实验流程见图1。
Figure 1. The basic process of soil microbial DNA sequencing
图1. 土壤微生物DNA测序的基本流程
2.3.3. 测序结果处理
去除HiSeq平台测序数据中低质量的reads后利用重叠关系使用软件FLASH将满足拼接条件的成对reads组装成Tags [24]。去除嵌合体,将相似度在97%以上的tags聚类为一个OTU,将OTU中丰度最大的序列作为代表序列,通过RDP classifer (v2.2)软件将OTU代表序列与数据库比对进行物种注释,置信度阈值设置为0.8 [25],去除没有注释结果和注释为古菌的OTU,剩余的OTU用于后期分析。
2.4. 数据处理
主要使用Excel 2016对原始实验数据进行录入与计算,用SPSS 22.0进行方差分析与相关性分析,用Dunnetts’ T3进行多重比较,用Origin 2018软件进行作图。
3. 结果与分析
3.1. 不同植被样地土壤理化性质差异比较
不同植被调查样地土壤理化性质测定的结果见表2。
3.1.1. 土壤pH
由表2可知,以外来树种(湿地松、桉树)为优势种的植被样地,其土壤pH为酸性,以乡土树种(枫香、任豆、青檀、柏木)为优势种的植被,其土壤pH大部分为中性或弱酸性;其中编号为MDZSS和DCSAS两个样地均为强酸性土壤(pH < 4.5),编号为XYFS样地为弱酸性土壤(pH = 5.5~6.5),编号为YMRD、XJQT、JXBM三个样地为中性土壤(pH = 6.5~7.5)。种植桉树和湿地松会使土壤明显酸化;种植枫香时,虽然也会使土壤酸化,但酸化的程度没有种植桉树和湿地松明显;种植任豆、青檀、柏木三种乡土树种,则能使土壤的pH保持中性。
Table 2. Basic physical and chemical properties of soil in different vegetation survey plots
表2. 不同植被调查样地土壤的基本理化性质
注:同列不同小写字母表示差异显著(0.05水平)。
3.1.2. 土壤含水量
由表2可知,本研究的不同植被样地土壤含水量在1.97%~10.28%之间,以柏木、青檀等乡土树种为优势种的样地,其土壤含水量显著高于以外来树种为优势种的样地,不同植被样地土壤含水量高低排序为:JXBM > XJQT > YMRD > XYFS > DCSAS > MDZSS。由此可知,乡土树种对根际土壤水分的保持能力显著优于外来树种。
3.1.3. 土壤有机质含量
由表2可知,大部分乡土树种样地的土壤有机质含量高于外来树种样地,但乡土树种枫香样地与外来树种桉树和湿地松样地却无显著差异,不同植被样地土壤有机质含量高低排序为:JXBM > XJQT > YMRD > MDZSS > XYFS > DCSAS。土壤有机质主要来源于植物的凋落物及分泌物,部分来源于动物和微生物残体,是土壤肥力的重要指标之一 [26]。按照我国第二次土壤普查养分分级表 [27] (下文简称养分分级表),柏木、青檀、任豆三个样地的土壤有机质含量达到一级标准(>40 g/kg);枫香、湿地松样地的土壤有机质仅达到三级标准(>20~30 g/kg),与青檀、柏木及任豆样地相比,差了两个等级;桉树样地的土壤有机质含量最低,仅达四级标准(>10~20 g/kg)。
3.1.4. 土壤氮含量
由表2可知,不同植被土壤全氮含量在1.29~6.53 g/kg之间,其高低排序为:JXBM > XJQT > YMRD > XYFS > DCSAS > MDZSS,乡土树种土壤全氮含量均高于外来树种。
本研究的不同植被样地土壤速效氮含量为78.58~431.39 mg/kg。除枫香样地外,其余乡土树种样地土壤速效氮含量均高于外来树种,不同植被土壤速效氮含量高低排序为:JXBM > XJQT > YMRD > MDZSS > DCSAS > XYFS。
3.1.5. 土壤磷含量
由表2可知,土壤全磷含量在72.26~357.38 mg/kg之间,不同植被样地土壤全磷含量差异表现为:XJQT > YMRD > JXBM > XYFS > DCSAS > MDZSS,乡土树种土壤全磷含量高于外来树种。
不同调查样地土壤速效磷含量为2.40~4.94 mg/kg,其高低排序为:XJQT > DCSAS > YMRD > MDZSS > XYFS > JXBM。在所有样地中,青檀样地的速效磷含量最高。
3.1.6. 土壤钾含量
由表2可知:土壤全钾含量在4.74~8.76 g/kg之间,其含量高低排序为XYFS > YMRD > XJQT > JXBM > DCSAS > MDZSS;不同植被样地土壤全钾含量差异显著,其中,枫树样地土壤全钾含量最高,湿地松样地全钾含量最低,乡土树种土壤全钾含量均高于外来树种;不同植被样地土壤速效钾含量为11.83~81.77 mg/kg,其高低排序为XJQT > JXBM > YMRD > XYFS > DCSAS > MDZSS,乡土树种土壤速效钾含量均高于外来树种。
3.1.7. 土壤肥力综合评价
因为不同植被样地的土壤理化性质差异较大,因此,为了说明本研究的不同植被样地土壤肥力的差异,采用主成分分析法对6种不同植被样地土壤的理化指标进行综合分析 [28] [29] [30]。由表3可知,本研究中不同环境因子的KMO检验系数大于0.5,Bartlett的球形检验显示,本研究近似卡方值为416.99,显著性值为0.00 (P < 0.05),说明数据显著,因此该数据可进行因子分析 [31]。
Table 3. KMO and Bartlett test
表3. KMO和Bartlett检验
土壤pH、有机质等土壤理化因子组成主成分的方差贡献率及特征值见表4。由此可知,6个主成分已经包含全部的原有信息,其中前3个主成分特征值较大(大于1),能较好反应总体特征,为使各主成分明显区分,经旋转后,第1、2、3主成分方差贡献率分别由56.807%、20.635%、13.795%变为49.429%、26.637%、15.171%,前3个主成分累计贡献率91.237%,即前3个主成分进行分析时,仅丢失原有信息的8.763%,但91.237%的信息仍得以保留 [28]。
Table 4. Principal component eigenvalues and contribution rate
表4. 主成分特征值和贡献率
旋转后主成分对应的特征值见表5,在第1主成分特征值较大的有pH、有机质、含水量、速效氮、全氮,在第2主成分特征值较大的有pH、全磷、速效钾,在第3主成分特征值较大仅有全钾。
Table 5. The eigenvalues corresponding to the principal components after rotation
表5. 旋转后主成分对应的特征值
说明:旋转方法为具有Kaiser正规化的最大变异法。
计算不同植被样地土壤在各主成分上的得分,样地在各主成分上的得分相加即为综合得分,综合得分反映了土壤的肥力大小,其结果见表6。第1主成分中,排名前三的是柏木、青檀、任豆,它们的有机质、含水量等含量较高;第2主成分中,排名前三的是青檀、任豆、枫香,它们的全磷、速效钾含量较高;第3主成分中,排名前三的是枫香、任豆、柏木,它们的全钾含量较高。6种不同植被样地土壤肥力大小排序为:柏木 > 青檀 > 任豆 > 枫香 > 湿地松 > 桉树。相对而言,柏木、青檀、任豆样地为高肥力土壤样地,枫香、湿地松、桉树样地为低肥力土壤样地。
Table 6. Principal component score, comprehensive score and ranking
表6. 主成分得分、综合得分及排序
3.2. 不同植被样地的土壤细菌Tags、OTU差异比较
3.2.1. 细菌测序数据过滤及Tag连接
经PCR扩增后,不同植被样地的土壤细菌测序数据过滤结果见表7。由此可知,不同植被样地的土壤细菌原始测序数据Raw Data长度为44.73~45.79 Mbp。经过过滤后得到的高质量测序数据Clean Data均为37.5 Mbp,数据保留率81.90%~83.84%,因而能较真实反映土壤细菌群落结构。
Table 7. Filtering table of soil bacterial sequencing data of different vegetations
表7. 不同植被土壤细菌测序数据过滤表
对过滤后获得的Clean Data进行拼接,结果见表8。由此可知,不同植被样地土壤细菌测序的原始序列在各样地约为75,000条,拼接率均在99.5%以上,拼接后去除引物序列后共得到441,673条Tags。
3.2.2. 细菌OTU聚类
经过数据筛选,去除低质量的DNA序列片段,将序列片段拼接后进行OTU聚类,不同植被样地的土壤细菌OTU聚类结果见表9。由此可知,YMRD的OTU数量最多,达到4413;XYFS、JXBM、XJQT的OTU次之,也均达到3000以上;DCSAS、MDZSS的OTU数量较低,分别为1930和1511。YMRD和DCSAS的OTU数量相差高达2902,表明不同植被样地土壤的OTU数量存在较大差异,也表明不同植被样地的土壤细菌群落存在较大差异。
Table 9. Statistics of OTU clustering results of soil bacterial communities in different vegetations
表9. 不同植被土壤细菌群落OTU聚类结果统计
3.3. 不同水平上土壤细菌群落组成
3.3.1. 门水平
桂西北地区蕴藏着巨大的微生物资源,在门水平上,本研究的不同植被样地的土壤细菌物种丰度柱状图见图2,图中每种颜色代表一种细菌门类,颜色比例大小表示该细菌门类的物种相对丰度大小。从图2中可见,不同植被样地的土壤细菌物种组成相似,但所占比例不同。其中,DCSAS样地土壤共检测到细菌29门,JXBM样地土壤共检测到细菌38门,MDZSS样地土壤共检测到细菌27门,XJQT样地土壤共检测到细菌36门,XYFS样地土壤共检测到细菌40门,YMRD样地土壤共检测到细菌42门。
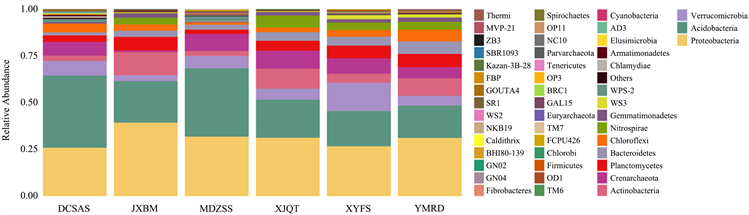
Figure 2. Histogram of species abundance of soil bacterial communities in different vegetation plots (phylum)
图2. 不同植被样地土壤细菌群落物种丰度柱状图(门)
Table 10. The relative abundance of soil bacteria at the phylum level
表10. 门水平土壤细菌相对丰度表
注:-表示该门类相对丰度低于1%或检测不到,样地所有相对丰度 ≤1%合并为稀有门类。
在本研究的不同植被样地土壤中,土壤细菌相对丰度大于1%的细菌菌群相对丰度总和分别为93.82%、96.88%、95.60%、98.45%、96.75%、97.22%,其具体分布见表10。其中,外来树种的土壤细菌优势菌均为酸杆菌门,次优势菌均为变形菌门,而乡土树种的土壤细菌优势菌为变形菌门,次优势菌为酸杆菌门,与之相反。在相对丰度大于1%的门类中,发现外来树种有WPS-2,乡土树种则没有;乡土树种有芽单胞菌门、硝化螺旋菌门,外来树种则没有发现这两个门类。在门水平上,外来树种土壤细菌物种组成及丰度上没有明显的差异;乡土树种样地中,枫香样地相对于柏木、青檀、任豆三个树种的样地,疣微菌门的相对丰度较高,而放线菌门的相对丰度较低。
OTU聚类结果显示不同植被样地土壤间OTU数量存在巨大差异,表明不同样地土壤细菌物种差异较大,影响样品聚类,为消除该影响,遂对不同样地土壤细菌相对丰度进行标准化处理(进行以10为底的log转化)。图3为在门水平上不同样地土壤细菌丰度热图,图中纵向的聚类结果表示土壤细菌在不同样地间群落差异情况,如果两个样地之间距离越近,相连接的线段(枝长)越短,则说明两个样地之间土壤细菌的物种组成及丰度越相似。由图3可以看出YMRD、XYFS聚为一类,且枝长最短,说明YMRD、XYFS两个样地土壤细菌的组成种类及丰度最相似;DCSAS、MDZSS聚为另一类,说明其在细菌的组成种类及丰度上相似度高;DCSAS、MDZSS聚成的类别与YMRD、XYFS、JXBM、XJQT聚成的类别通过最长的枝相连,说明外来物种与乡土树种在土壤微生物物种组成的相似情况存在较大差异。横向聚类可以看出不同细菌种类在6个样地间丰度相似情况,相似度高则分在同一支上,如硝化螺旋菌门和芽单胞菌门就在同一支上,说明硝化螺旋菌门和芽单胞菌门在桂西北不同植被样地土壤中丰度情况最相似。

Figure 3. Species abundance heat map of soil bacterial communities in different places (phylum)
图3. 不同样地土壤细菌群落物种丰度热图(门)
3.3.2. 属水平
本研究的不同样地土壤细菌在属水平上物种丰度情况见图4。由此可知,在属水平上,DCSAS、JXBM、MDZSS、XJQT、XYFS、YMRD六个样地土壤细菌稀有属(相对丰度小于1%的细菌菌群)相对丰度总和分别为90.93%、93.77%、93.26%、84.13%、78.66%、90.66%,除XJQT、XYFS两个样地外,其余样地土壤细菌稀有属均达到90 %以上。在优势属(相对丰度大于等于1%)中,Candidatus-Solibacter在DCSAS、MDZSS两个样地相对丰度大小分别为3.08%、2.05%,Candidatus-Koribacter在DCSAS、MDZSS两个样地相对丰度大小分别为2.90%、3.03%,而在JXBM、XJQT、XYFS、YMRD四个样地中Candidatus-Solibacter、Candidatus-Koribacter相对丰度低于1%或检测不到;红游动菌属(Rhodoplanes)在各样地相对丰度大小为DCSAS (1.98%)、JXBM (3.33%)、MDZSS (1.66%)、XJQT (2.29%)、XYFS (3.78%)、YMRD (2.20%);氨氧化菌属(Candidatus-Nitrososphaera)在XJQT、XYFS、YMRD三个样地相对丰度分别为9.66%、8.20%、6.08%,在DCSAS、JXBM、MDZSS三个样地相对丰度低于1%或检测不到;DA101在JXBM、XJQT、XYFS、YMRD在四个样地相对丰度分别为1.18%、2.71%、9.36%、1.06%,在DCSAS、MDZSS三个样地相对丰度低于1%或检测不到;甾类菌属(Steroidobacter)在JXBM、XJQT相对丰度分别为1.73%、1.21%,其余样地相对丰度低于1%或检测不到;Candidatus-Xiphinematobacter在DCSAS样地相对丰度大小1.11%,其余样地相对丰度低于1%或检测不到。其中,Candidatus-Solibacter为DCSAS样地的优势属,红游动菌属为JXBM样地的优势属,Candidatus-Koribacter为MDZSS样地的优势属,氨氧化菌属为XJQT、YMRD两个样地的优势属,A101为XYFS样地的优势属。

Figure 4. Histogram of species abundance at genus level (species with species abundance less than 1% are merged into Others)
图4. 属水平物种丰度柱状图(物种丰度低于1%的物种合并成Others)
本研究的桂西北岩溶地区不同植被样地土壤细菌属水平上聚类结果见图5。由此可见,属水平上,不同乡土树种中YMRD、XYFS两个样地土壤细菌的组成种类及丰度最相似,JXBM与其他乡土树种在土壤细菌的组成种类及丰度差异最大;DCSAS和MDZSS两个样地在属水平上,连接的线段长于门水平上的聚类结果,也表明了DCSAS和MDZSS属水平差异大于门水平。此外,乡土树种与外来树种在属水平上,土壤细菌的组成种类及丰度相差也较大,表明外来树种可以影响当地土壤细菌的群落结构。

Figure 5. Species abundance heat map of soil bacterial communities in different vegetations (genus)
图5. 不同植被土壤细菌群落物种丰度热图(属)
3.3.3. 种水平
不同植被样地土壤细菌在种水平物种丰度情况见图6。由此可知,在种水平上,DCSAS、JXBM、MDZSS三个样地稀有种(相对丰度小于1%的细菌菌群,下同)相对丰度总和为100.00 %,无优势种(相对丰度大于1%的细菌菌群,下同);XJQT、XYFS、YMRD三个样地稀有种相对丰度总和分别为94.73%、92.31%、94.83%,优势种均为氨氧化菌属-SCA1170 (Candidatus-Nitrososphaera-SCA1170)
3.4. 土壤细菌丰富度及多样性比较
不同植被样地土壤细菌的多样性指数见表11。土壤细菌测序的深度通常用coverage指数衡量,coverage数值越高,表示样品中DNA序列被检测出来的越高,该指数越接近1,说明样品中的所有物种测序时已经基本覆盖。由表11可知,6个样品的coverage指数均大于0.97,最高达到了0.993,说明本次测序深度几乎覆盖样品中的所有土壤细菌,能较好的反映样品中土壤细菌的真实情况。土壤细菌群落的丰富度通常用chao指数衡量,chao指数越大,细菌丰富度也越大。不同样地土壤细菌丰富度chao指数由高到低排序为:YMRD > JXBM > XYFS > XJQT > DCSAS > MDZSS,外来树种土壤细菌丰富度均低于乡土树种。土壤细菌多样性通常用shannon指数衡量,shannon指数越小,则多样性越小。在所研究的样地中,任豆样地的土壤细菌多样性最高,其丰富的土壤细菌,表明任豆能够给土壤微生物提供更多元化的营养,满足不同微生物生命活动中对养分的不同需求。不同植被样地土壤细菌多样性指数大小排序为:YMRD > JXBM > XYFS > XJQT > DCSAS > MDZSS。在土壤细菌多样性中,任豆样地土壤细菌多样性最高,湿地松样地土壤细菌多样性最低,乡土树种土壤细菌多样性均高于外来树种。综上分析,乡土树种的土壤细菌丰度、多样性均大于桉树、湿地松两个引种的外来树种,说明乡土树种更有利于土壤细菌丰富度、多样性的提高。

Figure 6. Histogram of horizontal species abundance (species with species abundance less than 1% are merged into Others)
图6. 种水平物种丰度柱状图(物种丰度低于1%的物种合并成Others)
Table 11. Soil bacterial diversity of different vegetations (OTU level)
表11. 不同植被土壤细菌多样性(OTU水平)
3.5. 土壤细菌功能预测
3.5.1. 一级功能预测
将土壤细菌16S基因序列的种属信息导入数据库中,与数据库中已知基因功能的基因序列进行对比,从而获得该基因的功能预测结果。在本研究中,桂西北岩溶地区不同植被样地获得的一级生物代谢通路功能基因预测结果见图7。由图7可知,本研究中不同植被样地土壤微生物(细菌)一级生物代谢通路功能基因包括:稀有基因(丰度小于5%合并为稀有基因,5.87%~6.38%)、环境信息处理(11.46%~13.03%)、遗传信息处理(16.00%~16.77%)、代谢(50.81%~51.58%)以及未知的功能基因(13.59%~14.20%),每种功能基因在不同植被样地土壤中相对丰度基本一致,波动不大。生物运动需要能量,而能量的产生就在生物的新陈代谢中,因而微生物基因中约一半为代谢基因,才能保持正常的能量供应。

Figure 7. The function prediction results of the first-level functional layer of soil bacteria in different vegetations (abundance less than 5% merged into rare genes)
图7. 不同植被土壤细菌一级功能层的功能预测结果(丰度小于5%合并为稀有基因)
3.5.2. 二级功能预测
不同植被样地土壤细菌的二级功能预测结果见表12。在土壤细菌二级功能中,共检测到细胞外结构、细胞骨架等24种基因功能,其中乡土树种与外来树种土壤细菌基因在编码细胞外结构、细胞骨架等多个功能上基因含量波动不大,但在编码防御机制、碳水化合物运输和代谢、细胞壁/膜/包络生物发生三种基因含量差异较大(该基因在乡土树种平均含量与外来树种平均含量大于0.5%)。
3.6. 土壤养分与土壤微生物的相关性
3.6.1. 理化性质与土壤细菌多样性
桂西北岩溶地区不同环境因子(土壤养分)与土壤细菌丰度前十的种类(门) Spearman相关性分析结果见表13。由此可知,放线菌门与土壤pH、含水量等6种环境因子相关性较大,而变形菌门、硝化螺旋菌门及芽单胞菌门与一种或两种环境因子相关性较大;疣微菌门、泉古菌门等4种门类与环境因子均无显著相关性。在9种不同环境因子中,土壤pH与浮霉菌门、硝化螺旋菌门等5种细菌显著相关,速效钾、全氮等6种环境因子与一至两门细菌种类相关,而全钾与速效磷与土壤细菌无显著相关性。
3.6.2. 土壤肥力与微生物种类的相关性
不同土壤肥力样地的细菌种类相对丰度大小(相对丰度前十的门类)分析结果见表14。由此可知,桉树等低肥力样地土壤在放线菌门、芽单胞菌门两个门类的相对丰度显著低于柏木等高肥力土壤,而在拟杆菌门、酸杆菌门等门类上无显著差异。
Table 12. The function prediction results of the secondary functional layer of soil bacteria in different vegetations
表12. 不同植被土壤细菌二级功能层的功能预测结果
Table 13. Correlation analysis between soil environmental factors and soil bacterial species (phylum)
表13. 土壤环境因子与土壤细菌种类(门)的相关性分析
注:相关性在0.01级别(双尾)显著用**标出,在0.05级别(双尾)显著用*标出。用土壤细菌英文前三个字母表示该门类,具体细菌种类参见表10,如Pro表示变形菌门,下同。
Table 14. Soil fertility and relative abundance of soil bacterial species (phylum level)
表14. 土壤肥力与土壤细菌种类相对丰度(门水平)
注:同一行不同小写字母表示差异显著(P < 0.05)。
3.6.3. 土壤肥力与土壤细菌的相关性
土壤细菌从土壤生态系统中获取营养物质,营养物质的多寡决定着土壤细菌的分布,土壤肥力与土壤细菌不同种类的Spearman相关性分析见表15,由此可知,土壤肥力与疣微菌门呈显著负相关,与芽单胞菌门、放线菌门呈显著正相关,与绿弯菌门、泉古菌门等其他7个门类细菌无显著相关性。
Table 15. Correlation between soil fertility and soil bacterial species
表15. 土壤肥力与土壤细菌种类的相关性
4. 讨论
4.1. 细菌种类差异
本研究通过高通量测序和土壤理化分析相结合,讨论了乡土树种与外来树种在桂西北岩溶地区土壤细菌群落结构组成与土壤养分的差异,在门水平上,变形菌门和酸杆菌门在不同植被样地土壤细菌群落组成中占绝对优势。一些研究者发现,在种植香蕉 [32]、玉米 [33]、白桦 [34]、茶树 [35] 等植物的土壤细菌丰度中,变形菌门都比较高,因此推测变形菌门在各种植物土壤细菌群落的组成中占优势,可能是因为其生长速度较快 [36],从而在竞争中处于优势。本研究的柏木、青檀等土壤有机质含量较高的样地中,土壤细菌中的变形菌门丰度也较高,这与李晓娜等 [37] 在荒滩植被恢复的研究结果一致;酸杆菌门属寡营养嗜酸细菌,能够降解复杂的木质素与纤维素 [38],其对pH有一定的耐受性,在偏酸性土壤中丰度较高 [39]。本研究还发现,在桉树、湿地松这些酸性低肥力土壤样地中,酸杆菌门为优势菌,与前人研究结果一致。
在本研究中,变形菌门与酸杆菌门相对丰度比值中,DCSAS = 0.67、MDZSS = 0.87,土壤均为强酸性土壤,土壤肥力也较低;而在JXBM = 1.77、XJQT = 1.55、YMRD = 1.80、XYFS = 1.41,土壤为中性或微酸性土壤,土壤肥力也较高,因而,可以通过变形菌门与酸杆菌门相对丰度比值,初步确定土壤肥力(笔者拟定变形菌门与酸杆菌门相对丰度比值大于1.5为高肥力土壤,在1至1.5为中肥力土壤,低于1为底肥力土壤),但又因土壤微生物的生长受海拔、温度、气候、土壤类型以及人为活动等诸多因素影响,变形菌门与酸杆菌门相对丰度比值大小与土壤肥力关系有待进一步验证,同时其他非优势菌与土壤肥力的关系也有待进一步研究。
在属水平上,本研究的不同植被样地土壤中存在着大量的稀有属细菌,丰富的细菌库增加了土地的潜在价值。Candidatus-Solibacter、Candidatus-Koribacter,在桉树、湿地松两个强酸性土壤样地相对丰度较大,对强酸性土壤表现出较强的耐性;红游动菌属在黑暗好氧条件下能很好生长,本研究涉及的样地植被郁闭度较高,可为其提供黑暗好氧的条件,故而在各种植被土壤中均有分布有红游动菌属;氨氧化菌属最适pH值为7.0~8.5,能利用铵盐的氧化满足自身能量需求,是一种化能自养细菌,故而在青檀、枫香、任豆三个中性土壤样地相对丰度较高,在桉树、湿地松两个强酸性土壤样地相对丰度低于1%或检测不到,但其在柏木样地相对丰度低于1%或检测不到,其原因有待进一步研究。
左锐 [40] 在研究水质演化中发现放线菌门起主要作用,放线菌门与水质质量具有强相关性;张健飞 [41]、陈雪花 [42] 等在土壤养分研究中发现,放线菌门与土壤养分也具有较大的相关性。在本研究中,放线菌门与土壤pH、土壤含水量等6种不同环境因子相关性较大,与前人研究一致,鉴于放线菌门对土壤养分具有较大的相关性,因而可将放线菌门作为衡量土壤理化特性好坏的生物指标。
4.2. 土壤理化性质的差异
在土壤含水量方面,人工种植的湿地松及桉树,其土壤含水量显著低于乡土树种为优势种的样地,该研究结果与王学春的研究结果一致 [43]。以外来树种为优势种的样地土壤含水量较低,是由于桉树、湿地松等速生树种对水分需求大,造成根系周围土壤水分含量低,同时土壤有机质具有涵养水源的能力 [26],在桉树、湿地松样地土壤有机质含量低,因而土壤涵养的水分较少。另外,可能是由于大面积人工造林导致的森林覆盖增加,能显著促进陆面蒸散发,造成区域土壤水分下降 [44]。
土壤养分影响植物、微生物的生长发育,决定着土壤生态系统的结构和功能 [45]。根据综合比较分析,桂西北不同植被调查样地土壤肥力高低排序为:柏木 > 青檀 > 任豆 > 枫香 > 湿地松 > 桉树。由于本研究的样地大多为石山坡地,夏季丰沛的雨水冲刷,会造成部分土壤养分的流失 [46],而本研究样品的采集正值多雨的夏季,同时也是植物生长的旺季,土壤养分消耗较多,所以,测得的土壤肥力指标可能会有所降低。
4.3. 土壤细菌丰富度与多样性的差异
在本研究中,乡土树种土壤细菌群落丰富度、多样性均高于外来树种,土壤细菌聚类结果也表明,两者的土壤细菌群落结构存在明显差异,其中,任豆样地土壤细菌丰富度及多样性为最高,湿地松样地最低,这可能与植物对环境的适应性有关。由于乡土树种在与土壤细菌长期的协同进化过程中,能更好地为其周边的土壤细菌提供多元化营养物质和适宜的微环境,从而使土壤中细菌群落随之增多,但外来树种因种植密度、树种单一及植物化感作用等多种因素的影响,不能为土壤细菌提供多元化营养物质和适宜的微环境,所以其根际土壤微生物群落结构及多样性与乡土树种有较大差异。任豆属于豆科植物,能与固氮细菌通过共生关系将氮固定,并且任豆属于落叶乔木,叶片凋落后能为土壤增加养分,因而任豆样地土壤丰富度及多样性均高于其他物种。在土壤有机质含量中,青檀、柏木、任豆土壤有机质含量较高,分解之后能为微生物的生长提供更多元化的营养物质,Xun [47] 等研究发现,土壤中有机物质的积累,可以改善土壤细菌群落结构,增加土壤微生物生物量,本文的研究结果与其研究结果一致。
外来树种在生长发育过程中,通过与土壤生态系统复杂的相互关系,影响其生长区域的土壤理化性质及根际土壤微生物群落结构。研究发现,葱芥、地肤、苘麻等植物能够改变入侵地的土壤微生物群落结构,从而影响其他植物对养分的吸收,降低生物的多样性 [48]。与本研究的结果,与该研究的结果相似。
4.4. 土壤细菌功能预测的差异
由表12比较可知,在基因二级功能预测上,乡土树种与外来树种在编码防御机制、碳水化合物运输和代谢、细胞壁/膜/包络生物发生这三种基因含量差异较大。在编码这三种基因中,外来树种平均含量均高于乡土树种,编码防御机制、细胞壁/膜/包络生物发生这两种基因含量较高,可能是土壤细菌对外来树种导致的强酸性土壤的适应;DCSAS、MDZSS编码碳水化合物运输和代谢基因含量较高,可能是DCSAS、MDZSS两个样地土壤肥力较低,微生物为了维持正常生命活动,提高了碳水化合物运输和代谢基因的表达,才能保证足够的能量。
4.5. 桂西北岩溶地区植被恢复先锋树种的选择
从生态效益方面考虑,在桂西北地区岩溶地区植被恢复和石漠化治理中,应当把乡土树种作为生态恢复的先锋树种,本研究中,以柏木、青檀、任豆、枫香等乡土树种为优势种的样地,其土壤微生物多样性、pH值、土壤含水量、有机质等土壤肥力指标均优于外来种桉树、湿地松的样地。任豆、青檀、柏木等乡土树种耐贫瘠,生长速度快,根系发达,穿透能力强,且相互形成根网,具有很好的固土保水能力,对岩溶地区的特殊生境具有很好的适应性 [49] [50],可以涵养水源和有效减少山体滑波、泥石流等自然灾害的发生。因此,在建设壮美广西,共圆复兴梦想的伟大征程中,乡土树种在岩溶地区生态环境保护中的重要作用应引起人们的重视。
5. 结论
根据以上实验结果及分析讨论,可以初步得到以下结论:1) 不同植被对土壤养分及土壤微生物群落结构的影响不同,乡土树种的土壤肥力和土壤微生物多样性均高于外来树种,不同植被样地土壤肥力的高低排序为:柏木 > 青檀 > 任豆 > 枫香 > 湿地松 > 桉树;土壤细菌多样性的大小排序为:任豆 > 柏木 > 枫香 > 青檀 > 桉树 > 湿地松。2) 外来树种样地的土壤理化性质较差,土壤微生物可能通过增加表达编码防御机制的基因,从而主动适应环境。3) 综合考虑不同树种对岩溶地区土壤养分和土壤微生物群落结构的影响,可以将任豆、柏木、青檀等乡土树种作为桂西北岩溶地区植被恢复和石漠化治理的优良先锋树种。
基金项目
广西自然科学基金(桂科自0832273),桂西北岩溶地区石漠化综合防治实验室(校政发[2016] 91号),河池学院硕士专业学位建设基金课题(2016HJA007),广西高校大学生创新创业计划训练项目(201910605084)。
参考文献
NOTES
*通讯作者。