1. 引言
腹泻是世界上五岁以下儿童的第二大死因。每年,全球约有17亿例腹泻病,约有525,000名儿童死于该病。肠毒素大肠杆菌(ETEC)是儿童腹泻相关死亡的5大致病性病原菌之一 [1]。ETEC的传播与粪便通过受污染的食物或水经口传播有关 [1]。在非洲、亚洲和拉丁美洲的中低收入国家,ETEC的流行率很高 [1]。ETEC菌株通过产生热稳定(ST)、热不稳定(LT)肠毒素或两种毒素的组合 [2] 引起急性腹泻。
ST肠毒素是一种小的非免疫原性肽,在100℃时稳定 [3]。ST与内源性鸟苷和尿鸟苷具有相似的一级和三级结构 [4]。这允许ST与肠道刷缘的鸟苷酸环化酶C (GC-C)受体结合,鸟苷酸环化酶C是内源性配体鸟苷和尿鸟苷的受体 [5]。ST结合介导GC-C受体构象变化,导致细胞内信使环GMP (cGMP)增加 [6]。结果,肠道电解质平衡被破坏,因为钠和氯离子的吸收减少,碳酸氢盐和氯离子的分泌增加 [6]。
LT肠毒素是一种异六聚体毒素,由催化A亚基(LT-A)和五聚体B亚基(LT-B) [7] 组成。毒性A亚基由eltA基因编码,由A1和A2结构域组成,由一个二硫键连接 [8]。B亚基无毒,编码为eltB基因。B亚基与Vibrio cholerae [9] 生产的霍乱毒素(CT)的B亚基有大约80%的同源性。LT-B也与CT结合的受体相同 [7]。五个LT-B亚单位排列成五聚环并与肠上皮细胞表面的五个单唾液酸神经节苷脂受体(GM1)结合 [7]。结合到GM1后,LT全脂毒素通过内向网(ER)逆行内吞作用而内化。然后,LT-A的A1结构域被转移到细胞质中,随后激活膜结合GTPase的Gsa单位。Gsa单位的ADP-核糖基化永久激活腺苷酸环化酶产生cAMP。因此,升高的细胞内cAMP激活蛋白激酶A (PKA),抑制钠氢交换器(NHE2和NHE3)吸收钠离子,以增加囊性纤维化跨膜调节器(CFTR)分泌氯离子 [10]。cAMP水平的升高也会激活NF-kB和促ETEC粘附的有丝分裂原活化蛋白激酶(MAPK)信号通路 [11]。
不同的分子和生化技术已经被用来鉴定病原体的不同毒力因子。本研究采用逆转录聚合酶链反应(RT-PCR)和被动乳胶凝集试验检测不同培养物和样品中LT肠毒素产生的ETEC染色。
2. 材料和方法
2.1. 样品DNA制备
本研究共制备了4个DNA模板样品,4份样本均来源于临床区提供的腹泻儿童患者样本采集。对于每个样品,用接种环刮取大肠杆菌菌落,并在50 ml水中稀释。为了溶解细胞膜并从细菌中释放DNA,将细菌悬液在94℃下孵育10分钟。为了去除细胞碎片,将细菌裂解物在冰上孵育3分钟,并在台式微凝胶中4℃,10,000 rpm离心5分钟。收集含有DNA的上清液进行RT-PCR。
2.2. RT-PCR
RT-PCR使用最终浓度为0.05 u/ml的Taq DNA聚合酶、75 mM TrisHCl (pH 8.8)、20 mM (NH4)2SO4、1.5 mM MgCl2、0.01% (v/v) Tweenâ20、0.2 mM dNTPs、各400 nM的eltA基因特异性正向(5’-AGCATCGCCA TTATATGC-3’)和反向(5’-GAGAGGATAGTAACGCCG-3’)引物的PCR混合物进行 [12]。每25 ml RT-PCR反应用10 ml样品DNA模板。阳性(+ve)对照组,每25 ml 的RT-PCR反应中使用50 ng含eltA基因的质粒。而阴性对照组未使用模板。RT-PCR条件如下:DNA模板在94℃变性4 min,然后进行35个周期的94℃ 30 s;55℃ 30 s;72℃ 1 min重复,最后一次延长至72℃ 7 min,然后保持4℃,每个样品单独重复6次。
2.3. 凝胶电泳和分析
将RT-PCR产物装载到SyberSafe染色的1%琼脂糖凝胶上。在同一凝胶上运行一个1 kbp的带有标准DNA片段的DNA梯来估计扩增产物的大小。凝胶在80 V下运行。凝胶在透照仪上进行评估,并记录凝胶图像进行分析。
2.4. 胶乳凝集试验
用胶乳凝集法 [13] 检测四份标本上清液中热不稳定肠毒素的存在情况。对于每个样品,将100 ml培养上清液与100 ml GM1神经节苷脂包被乳胶珠混合。在室温下孵育2分钟后,在黑色背景下观察到凝集。用热不稳定肠毒素重组大肠杆菌培养上清作为阳性对照。以大肠杆菌菌株为培养上清液,以缺乏热不稳定肠毒素为阴性对照。每个样品分别重复21次进行乳胶凝集分析。
3. 结果
3.1. 检测阳性对照组经RT-PCR扩增的eltA DNA片段大小
生成标准曲线来估计eltA扩增子的大小。为了建立一个标准曲线,选择了一些标准DNA片段大小,并测量了它们在凝胶中迁移的距离(表1)。标准曲线(y = −0.4137x + 4.4568; R2 = 0.9979)是根据标准片段在琼脂糖凝胶槽(Well)中迁移的距离绘制的对数对(图1)。阳性对照组,扩增的eltA-DNA片段从槽(Well)中向下迁移3.70 cm,估计片段大小为843对碱基。

Table 1. Distance migrated by standard DNA fragments and the DNA fragment amplified by RT-PCR
表1. 标准DNA片段与RT-PCR扩增的DNA片段的距离迁移
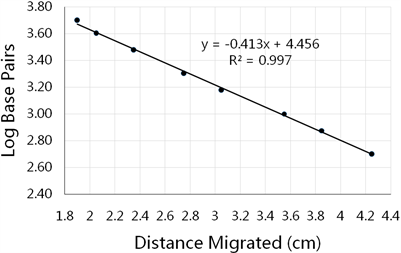
Figure 1. Log base pairs of standard DNA fragments vs distance migrated in the agarose gel. The linear line y = −0.4137x + 4.4568 was used to estimate the DNA fragment size amplified by RT-PCR; where, x = distance migrated by the DNA fragment and y = log10 (base pairs); and the linear regression coefficient of determination R2 = 0.9979
图1. 琼脂糖凝胶中标准DNA片段的对数碱基对与迁移距离的关系。用直线y = −0.4137x + 4.4568估计RT-PCR扩增的DNA片段大小,其中x = DNA片段迁移的距离,y = log10 (碱基对),测定的线性回归系数R2 = 0.9979
3.2. 用RT-PCR方法分析不同标本中eltA基因的表达
在样本1和样本3中,RT-PCR后只检测到一个片段大小,扩增片段DNA的大小在750~1000 bp之间。通过使用标准曲线,样本1和样本3中的扩增子大小为843个碱基对,与阳性对照中的扩增子大小相同(图2)。结果表明,样本1和样本3均表达eltA基因。而对于样本2和4,观察到多个大小的DNA片段(图2)。尽管RT-PCR后2号样本中发现了多个DNA片段大小,但2号样本中的DNA片段大小与阳性对照组中的DNA片段大小不符。因此,样本2没有eltA基因。然而,在样本4中,在不同的DNA片段大小中,有一条DNA带的大小与阳性对照相同。由于阴性对照中未发现843 bp的DNA片段,因此样品4中的DNA扩增不是由于PCR污染所致。虽然在样本4中发现了843 bp的DNA片段,但无法确定样本4是否含有eltA基因。这是因为PCR扩增后观察到多条带。
为了验证结果,对所有样品重复6次(表2)。在样本1中发现了一致的结果(表2)。对于样本2和样本4,大多数结果一致,在RT-PCR后观察到多条带。对于样本3,六次重复中有两次是不同的。在其中一个重复(Repeat 3)中,未检测到DNA片段;在另一个重复(Repeat 5)中观察到多条带(表2)。

Figure 2. Analysis of eltA gene expression in different samples as determined by RT-PCR. Each well was indicated by the product that was loaded. The Ladder contained DNA fragments with standard molecular weights. Positive control was indicated by +ve and negative control was indicated by −ve
图2. RT-PCR检测不同样本中eltA基因表达。每个条带上都显示了标准分子量的DNA片段。阳性对照用+ve表示,阴性对照用−ve表示
Table 2. Summarized results of eltA RT-PCR from 6 independent repeats
表2. 总结了6个独立重复序列的eltA RT-PCR结果
+表明eltA特异性DNA片段大小为843 bp,与阳性对照相同;−,表明RT-PCR后未检测到DNA片段大小;MB,多波段。
3.3. 被动胶乳凝集法检测不耐热肠毒素
对腹泻患儿上清液共4份进行凝集试验。乳胶凝集试验结果表明,样品3对GM1凝集呈阳性,而样品1、2和4对GM1凝集呈阴性(图3)。独立测试21次后,所有样品的结果一致。

Figure 3. A representative result of the analysis of the heat-labile enterotoxin with latex agglutination assay. Agglutination of the GM1 ganglioside-coated latex beads was evident by the clumps in the supernatants. Positive control was indicated by +ve and negative control was indicated by −ve
图3. 用胶乳凝集法分析热不稳定肠毒素的代表性结果。GM1神经节苷脂包被乳胶珠在上清液中的凝集明显。阳性对照用+ve表示,阴性对照用−ve表示
4. 讨论
在许多发展中国家,ETEC是引起急性腹泻的常见原因,尤其是儿童 [14]。确定和鉴定ETEC的毒力是非常重要的。本研究采用RT-PCR技术,结合毒力基因eltA的特异性引物,对产LT的ETEC菌株进行了鉴定。在两株大肠杆菌中发现了该毒力基因的存在,并证实这两株大肠杆菌是产LT的ETEC。尽管大多数结果在重复中是一致的,但在一些重复中观察到了阴性或阳性的故障。在处理样本和执行测试时,很可能是人为错误。在一些样本中发现多个扩增子大小,这很可能是由于引物设计。这个问题可以通过使用更特异的引物来解决。此外,多重PCR检测可在单一反应中检测和鉴别ETEC菌株和少数其他腹泻病原体 [4]。
用被动胶乳凝集法鉴定LT毒素的产生。在本研究中,有一个样本的乳胶凝集呈阳性,很可能是产生LT的ETCE。
在本研究中,产生LT的ETCE的患病率为25%。由于样本量较小,患病率可能较高。被动胶乳凝集试验方法简单、成本低,凝集反应结果一致、直观。然而,GM1神经节苷脂涂覆的乳胶珠可以与V. cholerae [7] 产生的霍乱毒素(CT)交叉反应。相反的被动胶乳凝集试验或使用特定的抗LT抗体可以提供更准确的结果来区分CT和LT毒素之间的差异。
综上所述,通过RT-PCR鉴定细菌中的毒力基因或分析样品中LT毒素的存在,可以鉴定热不稳定(LT)肠毒素产生的ETEC导致的腹泻。