1. 引言
20世纪中叶以来,随着工农业生产的发展、城镇化速度的不断加快和人类活动的日益活跃,重金属污染物被不断带入到土壤环境中,使得土壤重金属污染问题在世界范围内广泛存在且日益严重。2014年4月17日发布的《全国土壤污染状况调查公报》也显示,我国土壤无机重金属污染超标点位占到了全部污染超标点位的82.8%,已经成为了主要的土壤污染类型 [1]。污染土壤的重金属主要包括汞(Hg)、镉(Cd)、铅(Pb)、铬(Cr)和类金属砷(As)等生物毒性显著的元素,以及有一定毒性的锌(Zn)、铜(Cu)、镍(Ni)等元素。这些重金属污染物不能通过降解而从土壤中消除,因此当其在土壤中积累达到一定程度时,就会对土壤–植物系统产生毒害和破坏作用。重金属元素还能经由食物链富集和传递,严重危害农产品安全和人体健康。近年来,随着诸如2010年广西阳朔和2013年湖南“镉大米”等环境事件的曝光,重金属污染土壤的农产品安全问题开始受到我国民众的广泛关注。
在自然界中,重金属污染的形式是复杂多样的。单一金属元素构成的环境污染虽时有发生,但事实上并不存在绝对意义上的单一污染,通常重金属污染具有伴生性和综合性 [2]。土壤中的重金属污染往往是以某一种重金属元素为主,并同时伴随着其他元素的存在,即多金属并存的复合污染现象。如在对废蓄电池进行加工回收处理的周边土壤上,土壤中Pb的浓度高达12,000 mg/kg,同时还存在Cu和Zn严重超标的问题(1800~2200 mg/kg) [3]。这些重金属污染物之间相互作用,相互影响,对植物的毒害也呈现出多金属离子联合作用的复合效果,通常表现为不同金属之间的毒性协同、相加或拮抗作用。如与单独暴露于Cd/Zn污染土壤相比,在相同浓度的Cd-Zn污染土壤上种植的大豆(Glycine max)植株中,Zn的积累量会减少而Cd的积累量则会增加,植株生长迟缓 [4]。由此可见,与单一重金属污染相比,重金属复合污染中元素或化合物之间的交互作用显著影响着重金属污染物的生物效应(吸收、积累和毒性),对重金属复合污染下生物毒性效应的研究能够更客观地反映重金属生态环境风险。但由于土壤是一个复杂的有机无机多相体系,并且我国土壤类型众多,理化性质各异。这使得重金属元素之间的交互作用不仅会受到共存金属离子种类和浓度的影响,还会受到土壤理化性质和植物类型等多重因素的影响。因此,我们有必要开展基于我国典型土壤类型下重金属复合污染的生物毒性评价及其作用机制研究。
2. 重金属植物毒性评价模型的发展
2.1. 生物配体模型(BLM)
土壤重金属生物有效性/毒性评价模型是在水环境生物毒性模型的研究基础上建立起来的,并以单一金属的生物毒性评价作为主要研究对象展开。其发展主要经历了基于重金属总浓度的风险评价模型、自由离子活度模型(FIAM)、生物配体模型(BLM)和植物根细胞膜表面电势模型这四个阶段。它们之间既相互联系又彼此独立。BLM模型是在FIAM模型基础之上,考虑了环境介质如pH、共存阳离子(Ca2+、Mg2+等)等因素与金属阳离子之间的相互作用,主要是两者相互竞争生物体表面上的活性位点(BL),进而缓解金属阳离子的生物毒性。然而,BLM模型的“竞争”假说需要金属阳离子(如Cd2+、Cu2+)在生物体表面结合产生毒性效应的活性位点BL,同样也是共存阳离子(如Ca2+、H+)在生物体表面结合的位点,在此条件下产生竞争效应,缓解金属阳离子生物体毒性。有研究指出 [4] ,BLM模型“竞争毒性作用位点”的假设很难被证实,而且难以解释和评价大量阳离子(如Ca2+、Mg2+)增强金属阴离子(如
、
)的毒性现象。因此,尽管BLM模型已经成功的应用于多种金属元素和多种生物体对象,但其在合理的评价和阐述重金属生物毒性效应和作用机理上还存在不足。
2.2. 根细胞膜表面电势模型
基于生物体细胞膜表面电负性建立起来的植物根细胞膜表面电势模型,又称静电毒性/吸收模型(ETM/EUM) [5]。该模型的主要观点认为:在自然环境介质条件下,植物的细胞膜表面带有负电荷,使得植物细胞膜与土壤溶液之间形成一个双电层结构。这些负电荷主要来自于膜表面果胶羧酸、酸性氨基酸残基以及磷脂磷酸等基团的水解,其在细胞膜表面形成的电势就被称为细胞膜表面电势,用ψ0表示。研究表明,ψ0影响金属离子在本体介质(如溶液培养介质或土壤溶液)和膜表面的分布,从而对离子与植物相互作用过程有着重要影响,植物细胞壁的存在并不影响细胞质膜表面电负性ψ0。该模型示意图如图1所示,红色代表金属阳离子Mz+,绿色代表金属阳离子Mz−。ψ0通过静电作用增加金属阳离子在细胞膜表面的浓度,降低金属阴离子在细胞膜表面的浓度。当本体介质中的环境条件发生改变时,如溶液pH降低、大量阳离子含量(Ca2+、Mg2+等)增加时,这些阳离子又能够通过离子键结合(Ionic Binding)和电荷屏蔽(Charge Screening)的方式降低ψ0的负电性(即质膜表面去极化),如本体溶液中Ca2+从0.23 mM增加到2.50 mM,ψ0会从−45.6 mV增加到−24.4 mV,这会导致根膜表面富集的Cu2+的富集倍数减少36倍。由此可见,本体介质条件的改变会影响金属离子在细胞膜表面的富集浓度,缓解金属阳离子的植物毒性,加剧金属阴离子的植物毒性。目前,该模型已经被成功的应用于评价环境中大量阳离子(Ca2+、Mg2+等)缓解环境金属阳离子(Cu2+、Zn2+)的小麦根毒性,增强重金属阴离子(
、
)小麦根毒性的现象,并指出根表细胞膜表面电势的静电效应是产生金属毒性的重要作用机制之一。但静电作用机制并不否定专性竞争作用机制的存在。此外,有研究表明细胞膜表面的静电富集和排斥理论也能够较好的评价前人关于重金属植物毒性实验的数据 [6]。
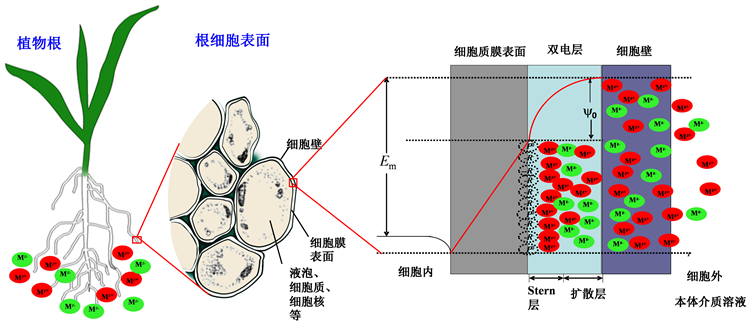
Figure 1. Schematic diagram of electrical profiles and ion distributions at a plant root cell surface, as described in the reference [7]
图1. 植物根细胞表面电荷特性和金属离子分配示意图,详细描述请参看文献 [7]
2.3. 生物毒性模型在重金属复合污染领域的应用
无论是渐趋成熟的BLM模型还是不断发展的细胞膜表面电势模型,它们都主要被应用于评价环境因素影响下的单一金属的生物毒性效应,而较少关注金属复合污染下的生物毒性研究。那么,能否将不断发展的评价单一金属毒性效应的模型引入到对金属复合污染下植物毒性的评价中去呢?模型的适用性及其在土壤介质上的应用性如何呢?
为了更好的为金属复合污染的环境风险管理提供指导,2012年在比利时的首都布鲁塞尔成立了一个国际性的研究小组—复合金属风险评估模型研究计划(MMME Research Project)。该研究计划提出模型建立的3个主要准则 [8] :1) 基于单一金属研究的生物有效性模型在被修正后可用于多金属复合污染的环境风险研究;2) 建立的复合污染评估模型要能够应用于评价不同金属组合形式,不同生物体对象,以及不同毒性效应终点下的实验数据;3) 已经建立的评估模型中要包括对金属形态的分析。基于以上要求,该研究组的学者们已经开始通过发展BLM模型来评价水环境中的金属复合污染的生物毒性效应,并取得了一定的进展。而在国内,陈中智等 [9] 也较早的将BLM模型引入到评价Cu-Pb复合污染下的绿藻生物体响应研究。由此可见,基于单一金属毒性风险评价的机制模型可以被引入到金属复合污染下的生物效应评价中,并成为完善金属复合污染环境风险评价的重要途径。
与对单一金属生物效应的评价相类似,BLM模型在阐明金属复合污染毒性作用机制方面还存在不足。如该模型“竞争毒性结合位点”假说能够很好的解释两种或两种以上金属的毒性拮抗效应(即各金属离子竞争相同的生物配体,一种金属降低另一种金属的生物毒性),也能够解释金属离子之间的毒性完全相加作用(即各金属离子存在各自独立的作用位点或生物配体,其毒性效应完全加和),但难以解释金属离子之间的毒性协同效应(即一种离子的存在能够增强另一种离子的生物毒性)。而细胞膜表面电势不仅能够更合理的界定金属的生物有效性(即膜表面的金属离子活度值),还能够在考虑离子质膜表面交互作用如竞争毒性结合位点的同时,从静电作用的角度为多金属复合污染的毒性评价提供一个新的视角,尤其是针对环境介质影响下的金属阴离子和金属阳离子复合污染毒性效应的研究。由此可见,我们有必要将细胞膜表面电势引入到金属复合污染植物毒性评价领域。调研文献结果也显示,目前只有不到10篇文献开展了这方面的研究,且主要集中于2010年之后。而基于我们前期研究也发现,在不同pH值条件下,细胞膜表面电势模型能够较好的评价Zn-Co复合污染下的小麦根毒性效应。
3. 土壤重金属的生物毒性作用机制
重金属元素在生物体亚细胞水平上的分布和结合形态也是产生生物毒性的重要机制,尤其是在土壤–植物系统中。Vijver在其发表的综述中指出,不同重金属在细胞内的作用靶位不同,仅分析元素在生物体组织中的总量是不够的,需进一步测定生物体各组织部分,如亚细胞组分的含量及分布等,从而深入研究和揭示重金属在生物体内的分布、代谢和毒性机制以及共存金属离子的影响机制 [10]。植物体内的亚细胞分布是操作定义上的区分,主要是通过连续差速离心将生物体分为重金属毒性敏感区(包括细胞器组分、细胞残渣部分和热应激蛋白组分)和重金属生物解毒区(包括微粒体组分和热稳定蛋白组分)。Li等 [11] 通过溶液培养实验的研究也证实与小麦根中Cd的总量相比,Cd在细胞残渣中的浓度能够更好的表征Cd的小麦根毒性。Wallace & Luoma [12] 研究指出重金属的亚细胞分布有助于理解重金属在生物体内的迁移风险,营养级有效金属(TAM)主要分布在生物体热应激蛋白组分、热稳定蛋白组分和细胞器组分中。此外,随着环境分子科学的迅速发展,特别是同步辐射技术的进步,为进一步解析重金属在细胞水平上的结合形态提供了充足的依据。Marie-Pierre Isaure等 [13] 通过基于同步辐射的微束X射线荧光影像技术(μ-XRF)来获得Cd在拟南芥根和叶组织中分布的热点区域(Hot Spot),并通过微束X射线近边结构光谱技术(μ-XANES)探明Cd在拟南芥根维管束组织中主要是与含S基团相结合,而Cd在叶表皮细胞中则主要是与含O/N的配体相结合,其研究结果能够让我们更好的理解Cd在植物体内的致毒和存储过程。
目前,亚细胞分离技术和同步辐射技术已经被越来越广泛的应用于重金属生物毒性机制的研究中,但其研究对象还大多集中于水生生物(如鱼类、藻类等)领域,或是基于模拟土壤溶液培养条件下的单一金属离子在陆生生物(如小麦、生菜等)体内的亚细胞分布和结合形态,较少关注土壤培养条件下复合重金属的植物体内研究。如Papazoglou [14] 研究发现Ni能够增加刺菜蓟(Cardoon)根中Cd的积累量,增加Cd从根中向地上部分的迁移能力。但该研究并没能从毒性作用机理上阐明共存金属离子是如何增加另一种金属的生物体内积累和迁移风险,而应用亚细胞分布技术和同步辐射技术来解析植株体内的重金属积累和赋存规律将会为解决这一问题提供新的途径,进而阐明重金属复合污染产生植物体毒性的内在机制。
4. 结语
近些年来,面对严重的土壤重金属污染问题,全国各地兴起了讨论和治理土壤重金属污染的热潮,广泛开展土壤重金属污染的防控工作。然而,我国现有的土壤重金属质量标准和生态风险评估工作仍然建立在重金属总量评估的基础之上,尚未考虑复杂的环境条件、共存金属离子对重金属生物有效性的影响。这将会加大土壤重金属修复工作的成本和难度。因此,我们亟需建立基于重金属尤是基于复合重金属污染条件下的生物毒性风险评估和生物体内作用机制的研究,以期能够服务于我国土壤重金属环境质量标准的修订和土壤重金属修复治理工作的实施。
基金项目
江苏省自然科学基金(BK20160859)。