1. 引言
森林是重要的碳储量基地,全球森林碳储量约为861 ± 66 Pg,其中44%储藏在森林土壤中 [1] 。森林转换是土地利用变化的重要方式,是影响土壤碳、氮循环的重要驱动因子 [2] [3] [4] 。过去几十年由于亚热带地区山高坡陡、土壤抗蚀能力差、降雨集中,亚热带丘陵区农业生产基地建设和综合开发高速发展,大面积天然阔叶林改为人工/经济林、果园和坡耕地。目前,我国人工林面积已达6933万公顷,占有林地面积的36.3% [5] 。森林转换导致植物地上部分生物量、凋落物等均发生明显变化,显著影响了土壤有机碳、氮的含量和转化过程 [6] 。Guo and Gifford (2002) [7] 总结了不同土地利用方式变化对土壤有机碳库的影响,表明天然林转换为针叶林,土壤有机碳储量平均下降了15%。森林转换导致的土壤有机碳数量和质量的变化使土壤有机碳的矿化过程发生显著改变。研究表明,云南松天然林和米槠天然林转换为人工林后土壤呼吸分别减少了25%和22% [8] [9] 。林型转换导致土壤不同形态氮素以及对土壤氮素转换过程起催化作用的脲酶活性都发生显著变化,从而对土壤氮矿化过程产生显著影响 [10] 。龚伟等(2011) [11] 的研究表明,天然常绿阔叶林人工更新成檫木林、柳杉林和水杉林后土壤全氮、铵态氮、硝态氮和微生物量氮含量和脲酶活性都显著下降。研究表明,亚热带常绿阔叶林转换为杉木林和竹林,土壤净氮矿化速率均显著下降 [12] 。本研究选取浙江省杭州市临安区玲珑山的天然阔叶林和由天然阔叶林转换而来的针阔混交人工林、马尾松人工林和杉木人工林为研究对象,采用室内培养法分析了亚热带天然林转换为人工林对土壤有机碳、氮矿化过程的影响及其机制,研究结果可为全球气候变化背景下森林土壤的碳、氮循环提供数据支持和理论依据。
2. 材料与方法
2.1. 研究区概况
研究区位于浙江省临安区西北部玲珑山,地理坐标为119˚39'E,30˚14'N,属于典型的亚热带季风气候区,该地区冬季温暖湿润,夏季气温较高,降水较多,四季分明,1月温度最低,7月温度最高,气温最低月份历年平均温度为3.4℃,气温最高月份历年平均温度为28.1℃,年平均气温15.8℃,年有效积温5774℃,年平均日照时数长达1939 h,年均降水量1613.9 mm,土壤为凝灰岩发育而来的黄红壤。研究区原为天然次生阔叶林,20世纪90年代初,一部分天然次生阔叶林改造为针阔混交人工林、马尾松人工林和杉木人工林,另一部分保留了原来的天然次生林。本试验选择坡度、坡向基本一致的天然阔叶林(BL)、针阔混交人工林(CB)、马尾松人工林(MP)和杉木人工林(CF)为研究对象,于2017年12月,在上述4种林地中,各建立20 m × 20 m的样地3个,在每个样地内采集4个0~20 cm土层的样品。将同一样地4个样品混合,作为该样地的土壤样品。4种林地基本情况如表1所示。
2.2. 研究方法
土壤有机碳矿化培养试验 [13] :称取50.0 g风干土放入500 mL的广口瓶中,调节土壤含水量至WHC的60%,在25℃恒温条件下培养15天。分别在第1,2,3,4,7,10,15天放入盛有10 mL 0.5 M NaOH溶液的小玻璃瓶用以吸收土壤排放的CO2,24 h后取出小玻璃瓶,用BaCl2溶液固定已被吸收的CO2,加入酚酞指示剂,用0.2 M HCl滴定至溶液由微红变为无色,记录HCl消耗量V,同时以不加土壤样品作为空白试验,记录HCl消耗量V0,计算有机碳的矿化量。
土壤净氮矿化速率 [14] :称取50.0 g风干土放入500 mL的广口瓶中,调节土壤含水量至WHC的60%,在25℃恒温条件下培养15天。分别在第1,2,4,10,15天测定土壤硝态氮(
-N)和铵态氮(
-N)含量。土壤
-N采用氯化钾提取–靛酚蓝比色法测定;土壤
-N采用氯化钾提取–紫外分光光度法测定 [15] 。
土壤有机碳采用重铬酸钾外加热法测定;土壤全氮采用半微量凯氏定氮法测定;土壤pH值采用水浸提酸度计法(土:水 = 1:5)。土壤蔗糖酶活性采用3,5-二硝基水杨酸比色法测定 [16] ;脲酶活性采用靛酚比色法 [17] ;过氧化氢酶采用高锰酸钾滴定法测定 [18] 。在每个试验样地进行地表(未分解和半分解层)凋落物的采集,整理后带回实验室,105℃杀青,70℃烘干,称取干物质的量,即为凋落物的现存量。

Table 1. The basic properties of the four different forest types
表1. 4种林地基本概况
注:BL:天然阔叶林Natural broad-leaved forests;CB:针阔混交人工林Mixed conifer and broadleaf plantations;MP:马尾松人工林Masson pine plantation;CF:杉木人工林Chinese fir plantation。下同。
2.3. 数据处理
土壤CO2释放量(mg kg−1) = 1/2(V − V0) × N × 44
式中,V:滴定处理过样品中NaOH溶液所用的标准HCl体积(mL);
V0:滴定空白实验中NaOH溶液所用的标准HCl体积(mL);
N:HCl溶液的物质的量(mol L−1);44为CO2摩尔质量(g mol−1)
净氮矿化量(mgkg−1) = (
-N + NO3-N)ti − (
-N + NO3-N)t1
式中,ti为培养后的天数;t1为培养第1天。
净氮矿化速率(mg kg−1d−1) = 净氮矿化量/培养天数
2.4. 数据统计
采用Microsoft Excel 2010和SPSS 18软件进行数据统计分析,图表中所有数据均为三次重复的平均值,采用单因素方差分析比较不同数据组间的差异,显著性水平设定为α = 0.05。
3. 结果与分析
3.1. 森林转换对土壤基本理化性质的影响
如表2所示,天然阔叶林转换为针阔混交人工林、马尾松人工林和杉木人工林,土壤有机碳含量显著减少(P < 0.05);全氮含量呈下降趋势,但处理之间差异不显著(P > 0.05);土壤pH没有显著变化。天然林转换为人工林,凋落物现存量显著减少(P < 0.05)。
3.2. 森林转换对土壤有机碳矿化的影响
如图1(a)所示,四种林分土壤有机碳的矿化速率呈现一致的变化规律,培养开始的1~4天CO2释放速率显著下降;从第4天至培养结束,CO2释放速率变化较平稳。培养期间,天然阔叶林、针阔混交人工林、马尾松人工林、杉木人工林土壤有机碳平均矿化速率分别为121.42、95.41、137.67和108.95 mg kg−1d−1,天然林转换为针阔混交人工林和杉木人工林土壤有机碳矿化速率显著降低,转化为马尾松人工林土壤有机碳矿化速率显著上升。
培养试验期间,天然阔叶林、针阔混交人工林、马尾松人工林和杉木人工林土壤CO2累计排放量分别为1250.64、1001.8、1404.14和1141.63 mg kg−1,天然阔叶林转换为针阔混交人工林和杉木人工林土壤CO2累计排放量显著降低,转换为马尾松人工林土壤CO2累计排放量显著升高(图1(b))。4种林地土壤有机碳矿化释放的CO2-C含量占总有机碳含量的比例分别为:1.62%、1.61%、2.98%和1.94%,天然阔叶林转换为针阔混交人工林和杉木人工林,其分配比例没有发生显著变化,转换为马尾松林,其分配比例显著增加(P < 0.05)。
Table 2. Soil physical and chemical properties of the four different forest types
表2. 4种林地土壤基本理化性质
注:同列不同字母表示不同林分之间差异显著(P < 0.05)。下同。

Figure 1. The dynamics of soil CO2 fluxes (a) and the cumulative CO2 (b) of different forest types during the incubation experiment
图1. 不同林分土壤CO2释放速率(a)和CO2累计释放量(b)的动态变化
3.3. 森林转换对土壤无机氮含量和净氮矿化速率的影响
如图2(a)所示,4种林地土壤
-N含量均随培养时间的增加而增加,至培养结束,4种林地土壤
-N含量顺序为:杉木人工林(123.57 mg kg−1) > 天然阔叶林(108.53 mg kg−1)>马尾松人工林(106.52 mg kg−1) > 针阔混交人工林(98.76 mg kg−1),杉木人工林土壤
-N含量显著高于其他3种林分(P < 0.05)。
如图2(b)所示,天然阔叶林、针阔混交人工林、马尾松人工林和杉木人工林土壤
-N含量分别由培养第1天的5.59、5.24、7.95和9.35 mg kg−1增加到了12.62、9.94、16.80和18.22 mg kg−1,至培养结束,杉木林土壤
-N含量最高,但处理之间没有显著差异。
土壤无机氮由
-N和
-N组成,其中
-N是矿化氮的主要构成部分,因此土壤
-N含量的变化对矿化氮含量具有重要影响。如图3(a)所示,在培养实验中,4种林地土壤无机氮含量均随培养时间增加而上升,至培养结束,4种林地土壤无机氮含量顺序为:杉木林(141.79 mg kg−1) > 马尾松人工林(123.32 mg kg−1) > 天然阔叶林(121.15 mg kg−1) > 针阔混交人工林(108.69 mg kg−1),杉木人工林土壤无机氮含量显著高于其他3种林分(P < 0.05)。
如图3(b)所示,培养期间杉木人工林土壤净氮矿化速率最高,为8.99 mg kg−1d−1,天然林转换为杉木林土壤净氮矿化速率升高;转换为针阔混交林和马尾松林,土壤净氮矿化速率呈下降趋势,但未达显著水平。
3.4. 森林转换对土壤蔗糖酶、脲酶和过氧化氢酶活性的影响
如表3所示,天然阔叶林转换为针阔混交人工林和杉木人工林,土壤蔗糖酶活性显著下降(P < 0.05);转换为马尾松人工林后土壤蔗糖酶活性显著增加(P < 0.05)。天然林转换为针阔混交人工林和杉木人工林土壤蔗糖酶活性分别下降了17.8%和66.7%,转换为马尾松人工林土壤蔗糖酶活性增加了71.1%。4种林地土壤脲酶活性存在显著差异,天然林转换为马尾松人工林和杉木人工林,土壤脲酶活性显著增加,分别增加了156.1%和429.6%;转换为针阔混交人工林,脲酶活性呈增加趋势,但未达显著水平。4种林地土壤过氧化氢酶活性没有显著差异。
如表4所示,4种林地土壤有机碳碳、氮矿化速率与酶活性的相关性分析的结果表明,蔗糖酶活性与有机碳矿化速率存在显著相关关系(P < 0.05),脲酶活性与土壤净N矿化速率存在极显著相关关系(P < 0.01),过氧化氢酶活性与土壤碳矿化速率和净氮矿化速率之间均无显著相关性(P > 0.05)。
4. 讨论
4.1. 森林转换对土壤有机碳矿化的影响
森林转换导致植物地上部分生物量、凋落物等均发生明显变化,显著影响了土壤有机碳、氮的含量和转化过程。本研究结果表明,天然林转换为针阔混交人工林和杉木人工林,土壤有机碳矿化速率显著降低,培养期间有机碳矿化释放的CO2量分别下降了19.9%和8.7%。天然林转换为人工林导致土壤有机碳矿化速率下降的原因主要有以下几个方面:1) 土壤有机碳含量下降 [19] 。本研究中,天然阔叶林转换为针阔混交人工林和杉木人工林,土壤有机碳含量分别降低了19.3%和24.0%,土壤有机碳作为有机碳矿化的底物,其含量降低导致有机碳矿化速率降低。2) 凋落物数量减少。本研究中,天然林转换为人工林后,凋落物数量均显著下降。盛浩等(2010) [20] 发现天然林改为人工林后,地上凋落物数量减少了13%~50%,土壤异养呼吸减少了24%~62%。3) 土壤酶活性的下降。土壤酶是有机质分解、周转和矿化的主要生物催化剂,也是土壤碳、氮循环的主要限制因素之一 [21] [22] [23] 。蔗糖酶是有机碳矿化过程中的重要酶,对土壤有机碳含量及其矿化过程有显著影响 [24] [25] 。本研究中,土壤有机碳矿化速率与土壤蔗糖酶活性之间存在显著相关关系,因此,天然林转换为针阔混交人工林和杉木人工林,土壤蔗糖酶活性的显著下降,是导致土壤有机碳矿化速率下降的原因之一。
本试验中,天然阔叶林转换为马尾松人工林,土壤有机碳含量和凋落物质量均显著下降,但培养期间土壤有机碳矿化CO2的累计释放量却提高了12.3%,这一方面可能是天然林转换为马尾松人工林后土壤蔗糖酶活性的显著升高所致。此外,土壤有机碳矿化释放的CO2-C分配比例是指在一定时间内,土壤有机碳矿化释放的CO2-C含量占土壤有机碳含量的比例,是土壤有机碳矿化速率的重要表征指标。土壤
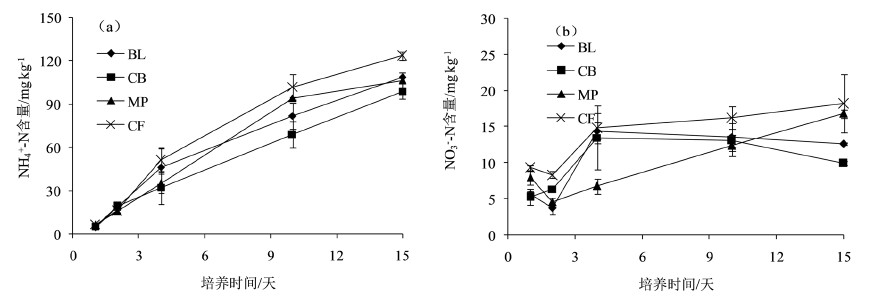
Figure 2. The dynamics of contents of soil
-N (a) and
-N (b) of different forest types during the incubation experiment
图2. 不同林分土壤
-N含量(a)和
-N含量(b)的动态变化
 不同字母表示不同林型之间差异显著(P < 0.05)。
不同字母表示不同林型之间差异显著(P < 0.05)。
Figure 3. The dynamics of contents of soil inorganic nitrogen (a) and net nitrogen mineralization rate (b) of different forest types during the incubation experiment
图3. 不同林分土壤无机氮含量的动态变化(a)和土壤氮净矿化速率(b)
Table 3. The soil en enzyme activities of the four different forest types
表3. 4种林地土壤酶活性
注b:同列不同字母表示不同林分之间差异显著(P < 0.05)。
Table 4. Correlations of enzyme activities with mineralization of SOC and nitrogen (n = 16)
表4. 土壤酶活性与碳、氮矿化速率的相关性(n = 16)
有机碳矿化释放的CO2-C分配比例从某种程度上反映了土壤的固碳能力,如果该比例越低,表明土壤的固碳能力越强,反之,则固碳能力越弱 [26] 。本研究中,天然阔叶林转换为马尾松人工林,土壤有机碳矿化释放的CO2-C分配比例显著增加,因此,天然阔叶林转换为马尾松人工林后土壤固碳能力减弱也是导致其矿化加剧的重要原因。
4.2. 森林转换对土壤氮矿化的影响
土壤中的氮主要以有机氮的形态存在,有机氮必须通过矿化作用、硝化反硝化作用等过程转化为能被植物吸收利用的有效氮形态。氮矿化是土壤氮素转化过程中的重要环节 [14] 。本研究中,天然阔叶林转换为针阔混交人工林和马尾松人工林,土壤净氮矿化速率没有发生显著改变,转换为杉木人工林,土壤净氮矿化速率显著升高。虽然土壤酶与土壤肥力之间不存在显著相关关系,但土壤酶能够促进土壤中物质转化与能量交换是不争的事实 [27] 。脲酶的酶促反应产物氨是植物氮源之一,它的活性可以用来表示土壤氮素状况 [28] [29] 。本研究中,4种林分土壤净N矿化速率与土壤脲酶活性显著相关,杉木人工林土壤脲酶活性显著高于其他3种林分,这与杉木人工林土壤净氮矿化速率显著高于其他3种林分的结果一致,表明土壤脲酶活性的显著增加可能是促进杉木人工林土壤氮矿化加速的重要原因。但本研究中,天然阔叶林转换为针阔混交人工林和马尾松人工林,土壤脲酶活性同样显著增加,却并未使2种林分的土壤净氮矿化速率得到显著提高,这表明森林转换导致的土壤净氮矿化速率的变化是一个复杂的生物过程,脲酶变化仅仅是其中的影响因素之一。
5. 结论
1) 天然阔叶林转换为针阔混交人工林和杉木人工林,土壤有机碳矿化速率显著降低;天然林转换为马尾松人工林,土壤有机碳矿化速率显著升高,土壤有机碳矿化释放的CO2-C含量占总有机碳含量的比例显著增加,土壤固碳能力减弱。
2) 天然阔叶林转换为针阔混交人工林和马尾松人工林,土壤净氮矿化速率没有显著变化;转换为杉木林,土壤净氮矿化速率显著增加,表明天然阔叶林转换为杉木人工林,土壤供氮能力显著增强。
3) 天然阔叶林转换为针阔混交人工林和杉木人工林蔗糖酶活性显著下降,转换为马尾松人工林蔗糖酶活性显著升高,土壤蔗糖酶和土壤有机碳矿化速率之间呈显著正相关关系。天然阔叶林转换为人工林土壤脲酶活性均呈增加趋势,杉木人工林土壤脲酶活性显著高于其他3种林地,土壤脲酶活性与净氮矿化速率呈显著正相关关系。
基金项目
浙江农林大学大学生创新项目(2013200040)。