1. 引言
小麦是世界上主要粮食作物之一,在我国小麦是仅次于水稻的主要粮食作物,其种植面积约为全国粮食种植面积的20%~27%左右,消费量占粮食总消费量的23%左右。种子引发(Seed priming)也称渗透调节(Osmotic conditioning),是使种子缓慢吸水的过程 [1] ,其原理是控制种子的吸水速率和最终的吸水量,允许种子内预发芽的代谢(但是要防止胚根的伸长) [2] [3] 。最简单的引发是水引发,是在控制给水条件下使种子定量吸水,达到促进萌发但并不引起吸胀伤害的吸湿回干技术 [4] 。种子引发能够使细胞膜结构重新排列,诱导一些有关抗性物质的合成 [5] [6] [7] ,使种子的细胞器、膜结构 [8] 、遗传物质、激素水平和相关酶类 [9] 处在种子萌发的最佳状态,从而加快种子的萌发速度,提高种子萌发的整齐度和对萌发时不良环境的耐性 [10] [11] 。大批量的种子引发技术含量较高,要严格控制引发时间,控制不当会导致过度引发,胚根突破种皮,甚至胚芽也伸长。不同活力的种子,引发时间存在一定差异,活力低的种子发芽慢,需要较长的发芽时间,而活力高的种子则发芽迅速,实验表明,小麦种子引发24 h胚根就可以突破种皮。
种子吸水起始一系列导致萌发的代谢事件,并使种子的脱水耐性随萌发进程逐渐丧失。在萌发的早期阶段和胚根突破种皮之前,大多数植物的种子能够干燥到它原来的含水量,而不引起伤害 [12] 。
Seraratna [13] 认为萌发36 h的大豆种子不能耐受脱水,吸胀40 h的豌豆种子脱水18 h后,幼苗不能耐受5%的含水量 [14] ;豌豆吸胀18~24 h,玉米吸胀48 h丧失脱水耐性 [15] ,从而可以看出,豆类种子萌发后普遍不耐脱水,而禾本科植物的种子研究较少,胚根突破种皮后多久丧失脱水耐性还有待研究。
为了探索过度引发给种子带来的风险,探索种子失水后的萌发潜力,本研究中设置将种子引发到正常状态(胚根未突破种皮)作为对照,将过度引发,即生物学上的萌发以后的不同状态(胚根出现、胚芽出现、胚芽继续生长)设为实验组,通过多种实验来探索种子脱水耐性的维持时间,不同状态下种子回干后二次萌发的萌发率,萌发后的状态等指标,以此来为商业化、规模化的引发及引发风险评估提供理论参考。
2. 材料与方法
2.1. 试验材料
小麦种子石麦22购自潍坊市种子销售点;发芽纸、发芽盒为本实验室提供。发芽试验采用托普300B型光照培养箱,种子贮藏采用恒温种子发芽箱。种子消毒采用1%次氯酸钠。
2.2. 种子引发处理
试验种子分两批进行处理,第一批在8月份(夏季高湿度)开始处理,种子为石麦22,浸种:将小麦种子浸泡在1%次氯酸钠溶液中5~8 min;将浸泡过的小麦种子用蒸馏水清洗3~5次;将清洗过的小麦种子浸泡在蒸馏水中,放在的恒温发芽箱中20℃吸胀4 h,之后将种子取出沥干,用湿纱布覆盖,20℃下,用塑料膜保湿,引发处理。将经引发的种子培养一定时间后分成两份,一份选取胚根突破种皮2 mm左右的,一份选取胚芽略大于1 mm的,于阴凉处平摊自然回干,回干至种子初始千粒重后开始贮藏试验。于20℃存放,分别存放0 d、2.5 d、5 d、7.5 d、10 d、20 d和30 d进行标准发芽试验,统计每天的发芽数。并于发芽试验的10 d、20 d统计仅有胚芽无胚根的种子数。
第二批于3月初(春季低湿度)开始,种子同为石麦22,消毒、浸泡吸胀等同上,种子分成四份,处理1引发10 h后回干;处理2胚根突破种皮2 mm左右胚芽未伸出时回干;处理3胚芽伸出1 mm左右,此时胚根约3~6 mm,回干;处理4胚芽3 mm左右,此时胚根约6~10 mm,回干,每份约3200粒,回干标准为回复到初始重量,每百粒4.46 ± 0.06 g。在发芽试验末期将幼苗去掉种子,120℃的烘箱中杀青30 min,然后在65℃的烘箱中烘干至衡重,记录干重。
2.3. 标准发芽及幼苗管理
透明塑料发芽盒,规格为13 × 13 × 10 cm,将其先用自来水冲洗干净,再用蒸馏水冲洗后烘干,用无水乙醇擦拭烘干发芽盒的内外表面。选择保水性良好的专用大张发芽纸,裁剪成16.5 × 12.5 cm的条状,置于烘箱内在120℃条件下烘干2~4 h,然后装入密封袋中待用。将发芽纸用蒸馏水浸透沥干后铺在发芽盒底,每盒两层,然后将引发到不同状态的小麦种子用镊子以10 × 10的方式摆在发芽纸上,第一批每组3次重复,第二批每组四次重复,置于智能光照培养箱中恒温20℃,8 h光照,16 h黑暗条件下发芽。
在标准发芽8 d时,记录发芽数,发芽期间始终保持适量的水分,避免水分过多引起的缺氧和水分过少产生的大量绒毛状气生根,发霉的种子冲洗后继续对其观察,发霉比例过高的更换发芽纸,腐烂种子淘汰,并随时计数。
2.4. 活力指数的计算
在标准发芽8 d过程中,每天记录发芽数,发芽结束后去掉种子,将幼苗在烘箱内105℃杀青2 h,70℃鼓风烘干至衡重,取出放干燥器降至室温,称重,以此作为幼苗干重,根据活力计算公式 [16] 计算种子的活力指数。
VI = ∑(Gt/Dt) × S
VI:活力指数;S幼苗的干重;Dt是发芽天数,Gt与Dt相对应的每天发芽种子数。
2.5. 数据分析与统计
通过SPSS2.0的一般线性模型(general linear model, GLM)进行方差分析,利用LSD法对各处理进行多重比较,检验本实验处理间的差异显著性。另外活力数据和干重数据利用SPSS2.0进行单因素方差分析,利用LSD法进行多重比较。
3. 结果与分析
3.1. 萌发后回干对种子发芽指标的影响
过度引发后种子是否耐受回干呢,如果耐受,种子在适宜的条件下应该可以继续生长呢?从统计结果分析看,回干对再萌发的影响的总体影响是显著的,贮藏和不同引发处理间的差异均达到极显著水平(表1)。
3.1.1. 两批贮藏种子的发芽指标
从图1可以看出,即使在夏季高湿度条件下贮藏,胚根突破种皮及胚芽大于l mm长的种子回干后,仍可以二次萌发,且发芽率在贮藏初期没有显著变化,仅从胚芽大于1 mm组在贮藏20 d以后开始有明显的发芽率下降趋势,而胚根刚出现就回干的种子贮藏30 d对发芽率没有显著影响。胚芽大于1 mm组发芽率最低下降至78%,贮藏15 d时为83%,说明贮藏对过度引发至胚芽伸长3 mm的种子有显著影响,长于20 d导致种子不能满足85%的发芽率基本要求,成为不合格种子。
春季贮藏虽然温度依然为20℃,但空气湿度和环境温度明显低于夏季,经过对春季贮藏种子的发芽率(图2)分析可知,贮藏15 d对胚根1 mm的处理3没有显著影响,而对处理4 (胚根3 mm)影响显著,到了30 d

Table 1. Analysis of variance between two factors of priming and storage
表1. 引发和贮藏两因素间的方差分析
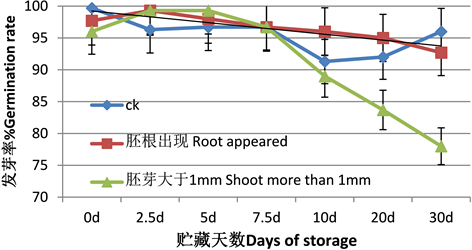
Figure 1. Changes of germination rate of seeds with different storage days under high humidity in summer
图1. 夏季高湿度下不同贮藏天数种子的发芽率变化

Figure 2. Changes of germination rate of seeds with different storage days under low humidity in spring
图2. 春季低湿度下不同贮藏天数种子的发芽率变化
二者发芽率都显著下降,但下降幅度较小,发芽率依然高于90%,在合格发芽率的范围内。胚根露出的处理2和未露胚根的处理1贮藏30 d并未显著降低其发芽率。说明,同样的贮藏温度下,环境湿度和温度对耐藏性有重要影响。春季低湿度与夏季高湿度相比,种子回干后存放发芽能力保持得更好。
3.2. 回干对二次萌发过程中胚根和胚芽的影响
由图3可看出,萌发2~5 d内,如图中A、C、E所示,正常的对照种子应该先长根,再长芽,胚根2~3 mm以上时芽才开始萌动,芽3 mm以上时,有数条种子根已经开始旺盛生长,而根露出以后再回干的种子,图3B为处理种子,二次萌发时大部分先长芽,再长根,大多数种子都如此,说明种子根对脱水敏感,从实验调查可知脱水后露出的根都死亡了,需要重新从根原基再发新根。
从表2可以看出,在二次萌发过程中,过度引发的种子常表现为先发芽后长根的现象,在发芽初期尤其明显,萌发第一天就有51%~69%的比例,到后期由于新根长出,渐渐恢复成正常幼苗,无根的幼苗因此随着萌发天数增加数量逐渐减少,到最后仍有少量根不正常的畸形苗存在。根回干后几乎不能在原处继续生长,说明根对脱水敏感,包在种皮内的部分还有再生能力和耐脱水性,随着过度引发的程度加
 注:图中A、C、E为正常干种子萌发的照片;B、D、F为萌发后回干的处理种子再次萌发的照片。
注:图中A、C、E为正常干种子萌发的照片;B、D、F为萌发后回干的处理种子再次萌发的照片。
Figure 3. Seed germination photos in different storage days under low humidity in spring
图3. 春季低湿度下不同贮藏天数种子的发芽情况
Table 2. The number of germination seeds with only shoots after storage 20 d under high humidity in summer
表2. 夏季高湿度下贮藏第20 d种子萌发过程中仅长胚根的种子个数
深,存储天数的增加,这种能力也随之降低。
萌发后回干,种子可耐受脱水,伸长3 mm的胚芽也表现了一定比例的脱水耐性,但也有畸形苗出现(图4),个别种子胚芽鞘尖端有少部分死亡,有些种子叶片卷曲或芽鞘开裂,大部分芽组织回干后依然可以继续正常生长,处理3和处理4萌发后杂菌感染较为严重。畸形苗的数量也持续增加,一部分畸形苗是因为胚根死亡,导致无根,少部分是胚芽或胚芽鞘的异常。由图5看出干种子和正常引发的处理1种子,畸形苗数量少,处理3和处理4种子随着存储时间的增加,畸形苗率增加显著,说明胚根和胚芽的损伤和丧失是渐变过程。
3.3. 引发程度与存储天数对活力指数的影响
那么引发和过度引发的种子存放不同时间后其活力的变化又是怎样呢,从图6可以看出,随着存储

Figure 4. Percentage of abnormal seedlings after seed germination
图4. 种子萌发后的畸形苗百分率
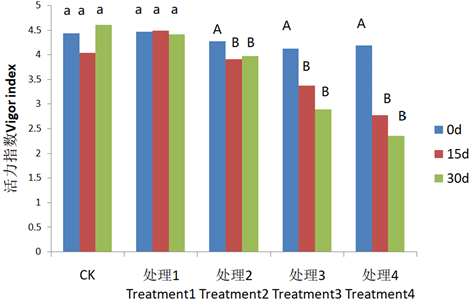 注:图中字母是经过单因素方差分析和多重比较后各处理间的差异显著性,小写字母显著性水平为P < 0.05水平显著,大写字母为P < 0.01水平显著。
注:图中字母是经过单因素方差分析和多重比较后各处理间的差异显著性,小写字母显著性水平为P < 0.05水平显著,大写字母为P < 0.01水平显著。
Figure 5. Changes of seed vigor index in different treatments under low humidity in spring
图5. 春季低湿度下各处理种子活力指数的变化
 注:图中字母是经过单因素方差分析和多重比较后各处理间的差异显著性,不同小写字母显著性水平为0.05,不同大写字母为0.01水平显著。
注:图中字母是经过单因素方差分析和多重比较后各处理间的差异显著性,不同小写字母显著性水平为0.05,不同大写字母为0.01水平显著。
Figure 6. Changes in dry weight of seedlings treated at low humidity in spring
图6. 春季低湿度下各处理幼苗干重的变化

Figure 7. Seedling growth on last days of germiantion after 0 d, 15 d and 30 d storage in spring under low humidity
图7. 春季低湿度下回干后存储0 d、15 d、30 d的幼苗生长情况
天数的增加,正常和处理1种子活力指数保持较高;而且引发可以提高种子活力,处理2种子活力指数下降达到显著水平;处理3和处理4活力指与对照相比极显著下降。随着引发、萌发时间的延长和存放天数的增加,活力指数下降速度也依次增大。说明种子脱水耐性下降的同时,种子的活力也在下降。
由图7看出对照种子和处理1种子生长量差异不显著,并且在30 d存储时间内基本保持稳定;处理1种子存贮0 d与对照相比生长量没有显著下降,存储0~30 d内生长量未见显著变化,说明处理1较适应短期贮藏;处理2、3、4种子未存储时生长量与处理2相当,略低于对照,说明即使不存储,回干本身对种子也有轻微影响,处理3及处理4则随着存储时间的增加再萌发后生长量降低显著。
由图6可以进一步佐证生长量的观点。从图7发芽末期幼苗状态照片可以看出,在30 d的存储时间内,对照、处理1和处理2种子能耐短期室温存储,表现了较高的脱水耐性,处理1、处理2长势要好于对照,处理3和处理4回干后,其生长量则随着存储时间的延长逐渐变差。说明胚芽出现后再回干会对幼苗生长量产生一定影响,再进行贮藏则会显著影响生长,从幼苗萌发及生长情况看过度引发后再在适宜条件下短期贮藏不会引起种子大量死亡。
4. 讨论
4.1. 贮藏对引发种子的影响
合理的引发应该在胚根突破种皮之前开始回干。在种子发芽过程中,胚根突破种皮之前,种子的活性增强 [17] ,各项生理代谢活动加强,为种子萌发做前期准备,萌发尚未启动,首次细胞分裂并未开始。有报导认为引发过的种子,具有高的种子活力,出苗迅速整齐,耐受性加强 [10] ,抵御外界不良环境的能力增强。引发是吸胀后再干燥的过程,在回干过程中种子重新积聚了大量耐受干燥的蛋白或小分子。此时各种酶的活性都已启动 [11] [18] ,生命活动依然比干种子旺盛得多,因此在较高的温度和湿度下种子内各种消耗加剧,贮藏时间越长,消耗越大,越不利于活力的维持。本研究引发后存贮初期,种子还能保持较高的活力,但时间越长,细胞就越不具备优势,本已经具备的引发优势也渐渐消失,直至活力下降到不及对照种子。
种子引发后贮藏的过程中,不同物种或者品种的生理变化过程并不一致,受贮藏时各种环境因子的影响较大 [4] 。引发时间越长,萌发的启动准备越充分,细胞活动能力越活跃,一旦胚根突破种皮,完成生理萌发过程,种子更难以降低生命活动,因此萌发后的可贮藏期会更短,活力下降也越快。本研究认为在常温高湿度条件下,种子可以贮藏10 d,维持较高活力,1个月明显降低活力,多数种子仍能长成正常幼苗。但发芽率显著下降;越晚停止引发种子的可贮藏性越差。常温低湿度下,种子安全贮藏期可以更长,这与水稻种子引发后贮藏的变化趋势有相似的趋势 [19] [20] 。
4.2. 回干对过度引发种子脱水耐性和抗菌能力的影响
生物学上的萌发范围为胚根突破种皮2 mm以前,胚根突破种皮2 mm以后为萌发后事件。本研究处理2、3和4都是萌发后回干,即使芽3 mm左右,胚根5~7 mm时回干仍有大部分种子能再度萌发,萌发过程中已经伸长的根全部死亡,不过还能再生新根,并逐步长成正常幼苗,说明萌发后小麦种子仍保持一定的脱水耐性,伸长的根完全没有脱水耐性,芽比较耐受回干。伸长的根死亡后从种子内能再生新根,说明种子内部尚存在一定的保护脱水伤害的功能,使根原基存活并能从种子内发生出来。虽然我们将发芽的小麦回干到正常小麦的含水量的状态,但是它的萌发相关的激素水平 [21] 和生理活动可能仍保持较高水平 [9] ,湿度高的夏季比湿度低的春季对贮藏时间更敏感。在干燥季节,常温或较低温贮藏1个月为较安全的期限,大部分种子仍保持萌发能力和较高的生长速度。萌发后的种子再度回干并不能使小麦种子致死,但能迅速降低活力,加速种子的衰老速度。
处于不同发芽状态的种子在组织结构、生理状态和含水量方面与正常小麦种子差别较大。本实验处理2到处理4种子萌发过程中霉变率明显高于对照和处理1,说明萌发后种子再回干并不能提高对微生物的抗性,相反,抗性明显减弱。
处理1适宜引发处理与对照相比种子霉菌感染率下降,可能引发过程提高了种子的抗病能力,且种皮完整,养分的外泄少。种子的引发回干过程,可能通过干燥缺水过程诱导了抗干燥相关基因的表达,从而提高了抗性和对逆境有适应性,使种子能够更好耐受生物胁迫 [22] 。
4.3. 过度引发种子的幼苗判断标准
按照种子检验中正常幼苗的判断标准,应该是匀称健壮 [16] ,正常的萌发顺序是先胚根再胚芽,而本研究因回干导致了已萌发胚根的死亡,二次萌发100%都不是先出现胚根,说明不能以传统的方式来衡量幼苗是否正常,到发芽结束期所调查的大部分幼苗都达到正常幼苗的标准,宜在萌发结束时判断幼苗。种子萌发后回干,再度萌发所产生的畸形苗率2%~18%,大大高于正常种子和引发种子的畸形苗率,其原因之一是回干导致胚根损伤;二是少量种子胚芽畸形,可能是在脱水过程中受到局部伤害,导致芽不能正常生长。从畸形苗的表现可知,种子的胚芽存在较高的脱水耐性,但这种脱水耐性持续的时间较短,随着贮藏时间的延长,畸形率越来越高。
5. 小结
引发过度会导致胚根胚芽的出现,回干以后直接后果是已经伸长的根死亡,芽可以耐受一定时期的干燥贮藏用于周转运输和流通,但常温高湿度下不及时播种会逐渐降低种子的活力,并且导致种子对外界有害菌的抗性减弱,对植物产生不良影响,随着新组织的生长,这些不良反应逐渐减弱。
致谢
感谢山东省自然基金对本项目的支持,感谢胡畔在预备试验中所做的工作。
基金项目
山东省自然科学基金(ZR2014JL017)。
参考文献