1. 引言
非酒精脂肪肝(Nonalcoholic fatty liver disease, NAFLD)是一类以肝脏脂肪变性 ≥ 5%为特征的常见肝脏代谢综合疾病,在全球范围内的发病率约为25%,造成了严重的医疗和经济负担 [1]。流行病学和基础研究表明,调节脂质代谢紊乱对防治NAFLD具有重要意义 [2]。目前,具有脂质调节功效的天然产物成分因安全性高、疗效明显的优势已成为研究热点。
荷叶为临床中常用的中草药,已被列入药食同源的名录 [3]。现代医学广泛将荷叶应用于干预肥胖症和高脂血症 [4]。脂脉康胶囊、降脂宁颗粒、荷丹颗粒等荷叶复方制剂均具有一定的减肥降脂的疗效 [5] [6]。越来越多研究证明荷叶生物碱是荷叶中发挥调脂功效的主要活性物质,可抑制体外脂肪酶 [7]、抑制体外脂肪细胞增殖分化 [8] 和降低高脂膳食诱导动物模型的血脂水平,缓解肝脏损伤和代谢紊乱,改善肝脏脂肪性病变,对NAFLD具有一定的防治作用 [9] [10] [11] [12] [13]。例如,护肝清脂片是由荷叶、泽泻、山楂等六味中药组成,在临床上主要用于脂肪肝的防治,能明显改善非酒精性脂肪肝模型大鼠的肝脂数,降低血脂和增强机体抗氧化能力,其中荷叶碱含量为0.15 mg/g [14]。但目前荷叶生物碱对NAFLD防治相关研究集中在体内动物模型上,与动物模型相比,以脂肪酸诱导人肝癌细胞HepG2构建NAFLD体外细胞模型的方法具有造模操作简单、周期短和成功率高的优点,可模拟肝细胞脂质积聚,肝脏脂肪变性的特点 [15] [16] [17]。本实验通过油酸诱导HepG2建立NAFLD的体外细胞模型,研究荷叶生物碱提取物(LLE)对脂肪变性HepG2脂质积聚的影响并探讨可能机制,以期为荷叶防治NAFLD和相关产品的研发提供一定理论基础。
2. 材料与方法
2.1. 试剂与仪器
材料:人肝癌细胞HepG2,购于上海中科院细胞库;荷叶,采自浙江省建德市;RMPI1640培养基,美国Hyclone公司;胰蛋白酶、青霉素–链霉素混合液、RIPA细胞裂解液,Biosharp公司;油酸,分析纯,上海麦克林公司;四甲基偶氮唑蓝(MTT)、饱和油红O染色液,北京索莱宝科技有限公司;血清白蛋白(BSA),大连美伦生物技术有限公司;非诺贝特(fenofibrate, FF),浙江大学校医院;TG试剂盒,南京建成生物工程研究所;Trizol试剂、逆转录试剂盒Prime Script TM、实时荧光定量PCR试剂盒TB Green Premix Ex TaqTMII,TaKaRa公司;
仪器:CKX41GF型倒置显微镜,日本奥林巴斯有限公司;Eon酶标仪,美国伯腾仪器有限公司;25 cm2细胞培养瓶、96孔、6孔细胞培养板,美国Thermo Fisher Scientific公司;Thermo Scientific Nanodrop 2000微量核酸蛋白分析仪,赛默飞世尔科技(中国)有限公司
2.2. 方法
2.2.1. 荷叶生物碱提取物的制备 [7]
采用酸性乙醇超声法制备荷叶生物碱提取物(LLE):荷叶洗净后在70℃烘干,粉碎后过60目筛,用塑料袋密封包装,置于−20℃保藏备用。称取适量荷叶粉末,用75%酸性乙醇(pH = 5)作为浸提溶剂,料液比为1:43,提取温度为63℃,超声提取功率为410 W,提取20 min,减压抽滤除去滤渣,将滤液旋转蒸发至干后得荷叶醇提物。用1%盐酸水溶液将其溶解,加入1% NaoH溶液调整pH至11,再次减压过率得到滤液,用氯仿反复萃取,将得到的氯仿层旋蒸至干后得LLE。
2.2.2. 细胞培养
将HepG2细胞培养于含有10%胎牛血清、1%青霉素–链霉素混合液的DMEM高糖完全培养基中,置于37℃,5% CO2环境中培养。待细胞长至80%~90%时进行消化传代。
2.2.3. 脂肪变性HepG2细胞模型的建立
参考刘畅 [16] 的方法制备得到10 mmol/L的油酸造模储备液。用DMEM高糖培养基稀释储备液至终浓度为0、0.2、0.4、0.6、0.8、1.0、1.2 mmol/L的油OA诱导液,分别培养HepG2细胞24 h。用MTT法测定细胞增殖活性,油红O染色和胞内TG含量测定评价胞内脂质积聚情况,筛选建立脂肪变性HepG2的最佳油酸浓度。
2.2.4. LLE对脂肪变性HepG2HepG2细胞细胞增殖的影响
采用MTT法检测不同浓度的LLE对HepG2细胞造模前后细胞增殖的影响。设置对照组(只添加培养基)、油酸造模组(600 µmol/L)、油酸(600 µmol/L) + 不同浓度LLE组。
2.2.5. LLE对脂肪变性HepG2脂质积聚的影响
采用对脂肪变性HepG2细胞生长无影响的LLE浓度进行实验。设置对照组(CK)、油酸造模组(600 µmol/L)、油酸(600 µmol/L) + LLE组(6.14 µg/mL、12.28 µg/mL)和油酸(600 µmol/L) + 阳性药物FF组(3.61 µg/mL)。采用的油红O染色和的胞内TG含量测定方法评价LLE对脂肪变性HepG2细胞脂质积聚的影响。
2.2.6. MTT法测定细胞存活率
收集对数期的细胞,将细胞接种于96孔板中,细胞悬液浓度为5 × 104个/mL,5000个/孔,每孔加100 μL。待细胞贴壁后,按照油酸造模实验(2.2.3)或药物干预实验(2.2.4)设置分组培养24 h。结束后吸去培养基,避光加入MTT终浓度为0.5 mg/mL的培养基,37℃反应4 h。反应结束后取出96孔板,小心吸取孔内培养基,加入DMSO 150 μL,室温振摇10 min,待紫色结晶物充分溶解后,酶标仪570 nm下测定OD值。计算细胞存活率。存活率 = (A实验组 − A空白组)/(A对照组 − A空白组) × 100。
2.2.7. 油红O染色及TG含量测定
收集对数期的HepG2细胞,将细胞接种于6孔板中,细胞悬液浓度为2.5 × 105个/mL,5 × 105个/孔,每孔加2 mL,置于37℃,5% CO2环境中过夜培养。按照油酸造模实验(2.2.3)或药物干预实验(2.2.4)设置分组培养24 h。结束后吸去培养基,加入预热的PBS 1 mL清洗3次,每次2 min。加入4%多聚甲醛1 mL,37℃固定30 min。加入PBS 1mL清洗3次。60%异丙醇1 mL浸洗30 s,加入PBS清洗1次。避光条件下加入油红O工作液1 mL,室温避光放置30 min,PBS清洗2次。加入60%异丙醇1 mL进行分色20 s,重蒸水清洗2次。加入苏木素500 μL染色4 min,自来水洗3次。显微镜下观察油滴染色情况。细胞内TG含量按照试剂盒说明书检测
2.2.8. 实时荧光定量PCR测定mRNA表达
采用Trizol法提取HepG2细胞总RNA,使用Nanodrop2000仪器测定260/280 nm波长处吸光度,两者比值在1.8~2.1之间符合RNA纯度要求。按照逆转录试剂盒说明书先去除基因组DNA反应,后将RNA逆转录为cDNA于20 μL反应体系中。按照荧光定量试剂盒说明书进行实时荧光定量PCR步骤。采用QuantStudio3qRT-PCR仪进行PCR扩增。阶段一:预变性,95℃,30 s,1个循环;阶段二:PCR反应,95℃,5 s至60℃,34 s,40个循环。产物经熔解曲线和扩增曲线确认特异性,应用2−△△Ct法计算RNA相对表达量。各引物序列详见表1所示:

Table 1. Real-time fluorescence quantitative PCR primer sequences
表1. 实时荧光定量PCR引物序列
2.2.9. 数据处理
数据统计采用SPSS17.0和GraphPad Prism8统计学软件进行,采用均数±标准差表示,IC50计算采用GraphPad Prism8中的非线性回归最小二乘法拟合分析,多组间比较采用one-way ANOVA单因素方差分析,p < 0.05为差异具有统计学意义。
3. 结果与分析
3.1. 脂肪变性HepG2细胞模型构建
不同浓度的油酸诱导液处理HepG2细胞24 h后对细胞存活率和细胞内TG含量的影响如表2所示。由下表可知,浓度在400~600 µmol/L范围内可小幅度促进HepG2细胞生长。800 µmol/L的油酸造模液对HepG2细胞增殖产生抑制,存活率为83.05% (p < 0.05)。在0~1000 µmol/L范围内,HepG2细胞内TG含量随着油酸造模液浓度增加而上升,600 µmol/L油酸处理后HepG2细胞内TG含量约为对照组(不加油酸)的3倍(p < 0.05)。但是油酸浓度达到1000 µmol/L时,TG含量较800 µmol/L油酸处理无显著性变化。
Table 2. Effects of different concentrations of oleic acid on cell viability and TG content in HepG2
表2. 不同浓度油酸对HepG2细胞生长和TG含量的影响
同列数据后的不同小写字母表示在p < 0.05水平差异具有统计学意义。
采用油红O染色法观察胞内脂滴形成情况。如图1可知,无油酸处理的HepG2细胞边缘清晰,核膜完整,胞内未见红色脂滴。油酸造模液处理后,胞内出现红色脂滴。随着造模液浓度增加,HepG2细胞内的脂滴的大小和数量均显著增加。600 µmol/L的油酸造模液处理HepG2细胞发生明显的脂肪变性,与TG含量结果吻合。综合细胞存活率、TG含量和脂滴形成的结果,本实验确定以600 µmol/L油酸培养HepG2细胞24 h作为建立NAFLD体外细胞模型的方法。此建模方式与文献记载基本吻合 [18] [19]。

Figure 1. Effects of different concentrations of oleic acid on steatosis in HepG2 (×400)
图1. 不同浓度油酸对HepG2细胞内脂滴形成的影响(×400)
3.2. LLE对脂肪变性HepG2细胞生长的影响
LLE对脂质变性HepG2细胞存活率如图2所示。与空白组(不加任何处理)相比,油酸单独处理促进HepG2细胞生长,存活率为空白组的112.50% (p < 0.05)。当油酸与LLE共同处理后,细胞生长先受到促进,当LLE浓度为24.56 µg/mL,细胞存活率为空白组的90.96% (p < 0.05),细胞增殖开始受到抑制。随着浓度的增加,LLE对细胞的抑制作用逐渐增强。经过软件分析可知LLE对脂肪变性HepG2细胞的IC50为73.77 µg/mL。因此,选择对细胞增殖无影响的LLE浓度,即6.14 µg/mL、12.28 µg/mL作为研究脂肪变性HepG2细胞脂质积聚的给药浓度。
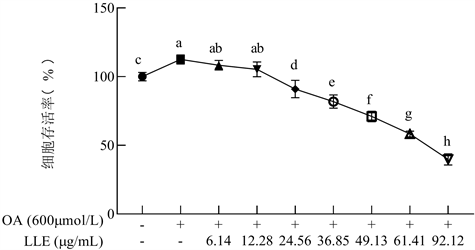
Figure 2. Effects of different concentrations of LLE on cell viability in HepG2
图2. 不同浓度LLE和对脂肪变性HepG2细胞存活率的影响
3.3. LLE对脂肪变性HepG2脂质积聚的影响
LLE对脂肪变性HepG2中TG含量的影响如图3所示。油酸造模后HepG2的TG含量为147.05 ± 16.82 µg/mg pro,约为CK组的2倍。浓度为6.14 µg/mL和12.28 µg/mL的LLE处理使TG含量分别下降20.35%和32.51% (p < 0.05),与阳性药物3.61 µg/mL的非诺贝特处理效果接近。由此可见,LLE可降低脂肪变性HepG2细胞中TG的含量,且高浓度作用更强。LLE对脂肪变性HepG2细胞内脂滴形成的影响如图4所示。观察可知,油酸造模组细胞周围出现明显的脂滴,密集环绕在细胞边缘。与油酸组相比,阳性药物处理后,脂滴明显减少。LLE处理组的脂滴密度减少,且高浓度的药物作用更加明显。油红O染色结果与TG含量检测结果基本一致。

Figure 3. Effects of different concentrations of LLE on TG content in OA cultivated HepG2
图3. 不同浓度LLE对脂肪变性HepG2细胞TG含量的影响
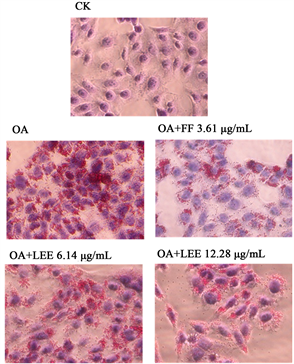
Figure 4. Effects of different concentrations of LLE on steatosis in OA cultivated HepG2 (×400)
图4. LLE对脂肪变性HepG2细胞内脂滴形成的影响(×400)
3.4. LLE对脂肪变性HepG2细胞脂质代谢相关基因mRNA表达影响
为进一步探究LLE降低脂肪变性HepG2细胞中脂质积聚的机制,采用实时荧光定量PCR检测高浓度LLE作用下脂肪变性HepG2细胞脂质代谢相关基因mRNA水平情况,结果如图5和图6所示。与CK组相比,油酸造模组的脂质合成相关基因SREBP-1c和ACCα的mRNA水平分别上调120.41% (p < 0.05)和70.87% (p < 0.05),脂质分解相关基因PPARα、CPT-1和ACOX1的mRNA水平分别下调40.64% (p > 0.05)、33.43% (p < 0.05)和39.19% (p < 0.05),由此可知脂肪变性HepG2细胞中脂质代谢相关基因表达紊乱。12.28 µg/mL的LLE处理组的SREBP-1c和ACCα的mRNA水平相较于油酸组无显著差异,但脂质分解基因PPARα、CPT-1和ACOX1的mRNA水平相较油酸组分别上调167.45% (p < 0.05)、76.21% (p < 0.05)和190.10% (p < 0.05)。由此可知,LLE可上调脂质变性HepG2细胞中脂质分解相关基因的表达,但对脂质合成相关基因的表达影响不大。
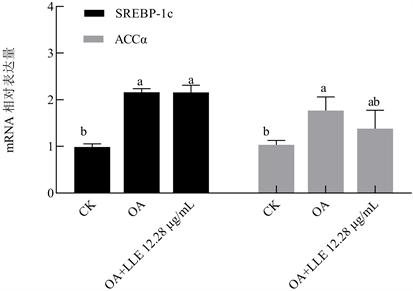
Figure 5. Effects of LLE on mRNA levels of lipogenesis related genes in oleic acid cultivated HepG2
图5. LLE对脂肪变性HepG2脂质合成相关基因mRNA水平的影响
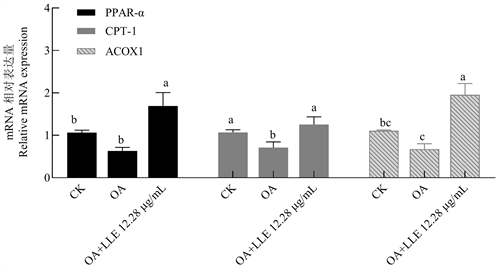
Figure 6. Effects of LLE on mRNA levels of lipidolysis related genes in oleic acid cultivated HepG2
图6. LLE对脂肪变性HepG2脂质分解相关基因mRNA水平的影响
4. 讨论
本研究基于脂肪变性HepG2细胞模型研究荷叶生物碱粗体物(LLE)对非酒精性脂肪肝脂质积聚的影响。LLE对脂肪变性HepG2细胞增殖的影响为低浓度促进,高浓度抑制,IC50为73.77 µg/mL。脂质积聚是肝细胞脂质代谢紊乱的表现 [20]。SREBP-1c是肝脏脂质合成的重要调控因子,参与TG的合成酶基因表达的调控,如ACCα、FAS等,其中ACCα是TG合成的关键限速酶 [21]。脂质的分解主要涉及脂质的转运和线粒体氧化的过程。CPT-1将长链脂酰油酸从胞质中转移到线粒体中,ACOX1主要涉及到脂肪酸的氧化及到ACOX1 [22]。PPARα是过氧化物酶体增生物激活受体家族中的一员,可抑制SREBP-1c从而影响FAS和CPT-1的表达,抑制TG的合成并促进脂质β氧化 [23]。本实验中LLE可降低脂肪变性HepG2细胞内TG含量和脂滴积聚程度,高浓度效果更为明显,进一步检测脂质代谢相关基因mRNA表达水平发现,LLE对TG合成相关基因SREBP-1c和ACCα的mRNA表达影响不大,但可显著增加TG分解相关基因PPARα、CPT-1和ACOX1的mRNA表达。相似的是,Zhang等人 [12] 发现荷叶碱单体通过上调PPARα/PGC1通路改善肝细胞的脂肪变性,促进肝脏脂肪β-氧化相关基因的表达,显著降低高血脂小鼠肝细胞中血脂水平。袁洋 [10] 通过SD大鼠非酒精性脂肪肝模型和代谢组学方法研究发现荷叶生物碱能调整造模后VLDL的合成和分泌障碍等脂质代谢紊乱,降低外周血中血脂含量,同时可以修复肝脏能量代谢紊乱,缓解脂质过氧化损伤和氧化应激,从而抑制大鼠炎症反应和肝脏脂肪性病变。
综上,荷叶生物碱提取物可抑制脂肪变性HepG2细胞增殖,降低细胞中TG含量和脂滴积聚作用,该作用与TG的β-氧化分解有关。本研究为荷叶生物碱防治NAFLD提供了一定的理论基础,同时对其在食品、保健品及药品的开发应用方面具有参考价值。
基金项目
药食同源功能健康食品开发——菊花、黄精及荷叶功能评价与健康食品开发。项目号:2021C02020。
NOTES
*通讯作者。