1. 引言
土壤微生物作为地球化学循环过程的重要参与者,参与土壤的有机质分解、元素循环和土壤矿化等化学活动,积极促进土壤物质循环、稳定与保持土壤的生产力和健康 [1] [2] 。近年来,土壤微生物多样性已成为土壤微生物学、土壤生态学和全球变化生物学等研究领域的热点问题,国内外众多学者对其开展了大量的研究 [3] [4] [5] 。
BIOLOG微平板法(BIOLOG-ECO Plate)是基于微生物利用碳源的表征特性来反映不同土壤或水体中的微生物区系差异,该方法能定量的评价生境来源微生物群落的功能多样性,具有方便快捷、灵敏度高、分辨率强、重现性好的优势 [6] [7] [8] 。虽然BIOLOG方法有一定的局限性,但其弥补了传统培养法、生物标记法和分子生物学方法无法获得的有关微生物群落总体活性和代谢功能信息的不足 [9] [10] 。目前被广泛应用于评价不同土壤类型、土地利用方式、土地经营管理模式、植被类型和植被恢复方式、海拔高度等的土壤微生物群落功能多样性 [11] - [16] 。因此,开展微生物群落研究具有非常重要的理论和应用价值。
鹦哥岭自然保护区位于海南岛中南部黎母山脉中段,属海洋性热带季风气候,区内保存着华南原生性最强、连片面积最大的雨林,是我国热带雨林生态系统保存最完整的区域之一 [17] 。鹦哥岭自然保护区具有海拔高、垂直带谱完整、生态类型丰富、原始性强等特点,目前的研究多集中于优势动植物的组成分布特征、多样性及其对环境变化的响应,以及土壤碳储量、凋落物现存量、养分循环动态以及森林生态系统服务价值评估等 [18] - [23] 。较少关注具有完整垂直谱带的热带山地雨林土壤微生物物种丰富度、群落组成结构、群落构建机制,以及与地上植被群落关系、与环境因子间关系的研究。为此,本文采用BIOLOG-ECO技术,通过沿鹦哥岭热带山地雨林海拔梯度的8个固定样地开展土壤微生物多样性研究,以期揭示其底物代谢多样性的变化特征及其土壤微生物适应海拔变化的生理代谢特征,为探索土壤微生物在热带山地雨林生态系统中的物质循环和能量流动中的作用和地位提供基础数据,为鹦哥岭自然保护区的管理提供理论依据。
2. 材料与方法
2.1. 研究区域概况
海南鹦哥岭国家级自然保护区(以下简称“保护区”),是以热带雨林及其生态系统为主要保护对象的森林生态类型保护区。保护区位于海南岛中南部黎母山脉中段,与霸王岭国家级自然保护区、佳西省级自然保护区交界,总面积50,464 hm2。保护区属热带海洋性季风气候;年均气温23~35℃,年降水量在1500~2000 mm之间,雨季旱季分明 [24] 。保护区土壤类型以赤红壤、砖红壤、山地黄壤土为主;地势中间高四周低,海拔从200~1812 m,最高峰为鹦哥岭 [25] 。“中高周低”的地貌特征形成了与海拔变化相应的土壤和植被类型,主要包括热带低地雨林、热带季雨林、热带山地雨林、热带山地常绿林、热带针叶林和热带山顶矮林等类型,整个区域的植被构成体现了完整的热带–亚热带植被垂直带 [26] 。
2.2. 样地设置与取样方法
2020年6月,选取保护区内不同海拔热带山地雨林的8个固定样地(50 × 50 m),其具体信息见表1。在每个固定样地内随机选取3个小样格(10 m × 10 m),即3个重复。在每个小样区内以五点采样法随机采样,先收集采样点30 cm × 30 cm范围内的凋落物装入自封袋,再采集新鲜表层土壤(0~10 cm),去除植物根系和石块等,将5个采样点的土壤充分混匀合并为一份样,装入无菌自封袋置于4℃保温箱里带回实验室,24 h内完成Biolog测定。

Table 1. Descriptions of the fixed sample plots at different altitudes in the tropical mountain rainforest of Yinggeling
表1. 鹦哥岭热带山地雨林不同海拔固定样方情况
注:1、YGL34表示34号固定样地;2、经、纬度数值系公里网格,采用北京54坐标系统。
2.3. 土壤、凋落物理化性质及酶活性测定
土壤理化指标的测定主要参照《土壤农业化学分析》和《森林土壤分析方法》 [27] [28] 。其中:土壤pH采用2.5:1水土比玻璃电极法测定,土壤含水量采用烘干称重法,土壤有机质采用高温外热重铬酸钾氧化–容量法,土壤全氮采用凯氏–蒸馏滴定法,土壤全磷采用氢氧化钠熔融–钼锑抗比色法,土壤全钾采用氢氧化钠熔融–火焰原子吸收分光光度法,土壤碱解氮采用碱解扩散法,土壤有效磷采用盐酸–氟化铵提取–钼锑抗比色法,土壤速效钾采用乙酸铵提取–火焰原子吸收分光光度法;土壤铵态氮采用硝态氮,氯化钾提取–氧化镁–代氏合净金蒸馏法,土壤硝态氮采用紫外分光光度校正因数法。
土壤酶活性的测定参照《土壤酶及其研究方法》 [29] 。其中:土壤脲酶采用靛酚蓝比色法,土壤酸性磷酸酶采用磷酸苯二钠比色法,土壤过氧化氢酶采用容量法。
凋落物于85℃烘箱烘干至恒重,以干物质计算凋落物生物量,并计算含水量。凋落物烘干后粉碎,按照LY/T 1271-1999《森林植物与森林枯枝落叶层全氮、磷、钾、钠、钙、镁的测定》测定凋落物的全氮、全磷和全钾养分指标。其中:凋落物有机碳采用高温外热重铬酸钾氧化–容量法;凋落物全氮采用硫酸–双氧水消煮–蒸馏滴定法;凋落物全磷采用硫酸–双氧水消煮–钒钼黄比色法;凋落物全钾采用硫酸–双氧水消煮–火焰原子吸收分光光度法。
2.4. 土壤微生物活性测定
参考章家恩等人的方法并加以改进,具体方法如下:称取鲜土10 g加入到装有90 mL 0.85% NaCl灭菌生理盐水的250 mL三角烧瓶中(瓶内加5~10粒玻璃珠),120 r/min振荡培养30 min,确保细菌、真菌及其孢子混合均匀。冰浴中静置5 min后,取5 mL土壤悬浊液加入到装入45 mL无菌水的100 mL三角烧瓶中,充分摇匀,重复稀释2次,得到1:100的土壤悬浊液用于ELSIA反应。用排枪吸取150 μl土壤悬浮液至已预热的Biolog-ECO平板的96孔中,28℃恒温条件下连续培养9 d,每24 h用Biolog自动读数仪读取590 nm和750 nm处的吸光值,记录数据并保存,用于后续分析 [30] 。
平均颜色变化率(Average Well Color Development, AWCD),也称平均吸光值,用来评估土壤微生物群落对碳源利用的整体情况及利用活性 [31] 。Shananon-Wiener指数(H’值),用来评估微生物物种的丰富度,指被利用的碳源的总数,为每孔中(C-R)值大于0.25的孔个数。对于稀疏种敏感,是研究群落微生物物种及其个体数与分布均匀程度的综合指标 [32] 。Simpson指数(D值),又称优势度指数。用来评估物种的优势度,该指数对富集中相对敏感,是对微生物多样性方面的集中性的度量。Pielou指数(E值),又称均匀度指数,是微生物群落实测多样性与最大多样性的比率,通过Shananon-Wiener指数计算出的精度。McIntosh指数(U值),是基于群落物种的多维空间上的Euclidian距离的多样性指数 [33] 。
(1) AWCD = [∑(Ci − R)]/n,式中:Ci为所测定的31种碳源孔的吸光值,R为对照孔的吸光值,n为培养基的碳源种类(ECO板为31种);
(2) H’ = −∑(Pi.lnPi),式中:Pi为第i孔相对吸光值与整个平板相对吸光值总和的比值,Pi = (Ci − R)/∑(Ci − R),表示有培养基(碳源)的孔和对照孔(A1)的光密度值差与整板总差的比值;
(3) D = 1 − ∑Pi2;
(4) E = H’/lnS,式中:S为被利用的碳源总数,log选用e为底;
(5) U = ∑ni2,式中:ni = Ci − R,是第i孔的相对吸光值,丰富度指数R指被利用的碳源总数目。
2.5. 数据处理与分析
所有数据均为平均值 ± 标准误,利用SPSS22.0 (IBM Corp., Chicago IL, USA)对各类碳源的利用及多样性数据进行单因素方差分析(one-way ANOVA)和主成分分析。Pearson相关系数来分析土壤及凋落物理化性质及酶活性与土壤微生物群落多样性指数之间的关系。
3. 结果与分析
3.1. 不同海拔土壤理化性质及酶活性特征
Table 2. Soil physical-chemical properties at different altitudes
表2. 不同海拔样地土壤理化性质
注:同列不同小写字母代表在各海拔梯度之间差异显著(p < 0.05)。
从表2可以看出,不同海拔样地土壤pH介于3.85~4.92之间,属于酸性土壤。南坡样地土壤pH随海拔的升高先升高后降低,1355 m海拔处样地的pH显著最低;北坡样地土壤pH随海拔的升高持续降低,458 m海拔处样地的pH显著最高(p < 0.05)。不同海拔样地土壤有机质含量29.43~116.80g.kg−1之间,整体随海拔的升高而升高,仅部分海拔样地间差异显著(p < 0.05)。不同海拔样地土壤全氮含量在1.02~4.15 g∙kg−1之间,整体随海拔的升高而升高,部分海拔样地间差异显著(p < 0.05);不同海拔样地土壤碱解氮、铵态氮和硝态氮含量随海拔的升高呈现不规则变化,仅部分海拔样地间差异显著(p < 0.05),碱解氮和硝态氮含量分别在550 m和1355 m海拔处显著最高(p < 0.05),而铵态氮含量在591 m海拔处显著最低(p < 0.05)。不同海拔样地土壤全磷和有效磷含量分别在0.12~0.40 g∙kg−1和1.31~4.44g∙kg−1之间,均随海拔的升高呈现不规则变化,且均在745 m海拔处显著最高(p < 0.05),部分海拔样地间差异显著(p < 0.05)。不同海拔样地土壤全钾和速效钾含量分别在6.38~32.19 g∙kg−1和81.16~168.37 g∙kg−1之间,均随海拔的升高呈现不规则变化,仅在部分海拔样地间差异显著(p < 0.05),全钾含量和速效钾含量分别在550 m和1025 m海拔处最高。
从表3可以看出,不同海拔样地土壤过氧化氢酶、酸性磷酸酶以及脲酶含量分别在2.50~3.64 mL∙g−1∙20min−1、1176.47~3067.77 mg∙g−1∙2h−1和460.79~858.86 mg∙g−1∙24h−1之间,均随海拔的升高呈现不规则变化,且仅在部分海拔样地间差异显著,其中土壤过氧化氢酶含量在1355 m海拔处显著最高(p < 0.05),在591 m海拔处显著最低(p < 0.05);土壤酸性磷酸酶和脲酶含量均在1025 m海拔处最高。
Table 3. Enzyme activities of soils at different altitudes
表3. 不同海拔样地土壤酶活性
注:同列不同小写字母代表在各海拔梯度之间差异显著(p < 0.05)。
从表4可以看出,不同海拔样地处的凋落物重量差异均不显著,凋落物其他理化性质均随海拔的升高呈现不规则变化,591 m海拔处样地凋落物有机碳含量显著最低(p < 0.05),其他理化性质仅在部分海拔样地间差异显著(p < 0.05)。
Table 4. Litter material physical-chemical properties at different altitudes
表4. 不同海拔样地凋落物理化性质
注:同列不同小写字母代表在各海拔梯度之间差异显著(p < 0.05)。
3.2. 不同海拔土壤微生物的平均颜色变化率的动态特征
由图1可知,24 h内土壤微生物活性较低,微生物培养24 h后,微生物对底物利用快速增加,各海拔样地AWCD值呈相似的趋势。整体来看,591 m海拔处的AWCD值最高,406 m海拔处的AWCD值最低,其他海拔处的AWCD值相当。由于不同类型的土壤微生物群落利用碳源能力的不同,因此不同的碳源利用模式即可表征微生物群落的差异。根据AWCD值曲线的变化情况(图1)还可以看出,南坡745 m海拔处样地土壤微生物个体生物量多且种群丰富,属于土壤微生物利用碳源的理论模式I;北坡800 m和南坡1025 m、1355 m海拔处样地土壤微生物个体数量少但种群丰富,属于土壤微生物利用碳源的理论模式II;南坡406 m、550 m和北坡591 m海拔处样地土壤微生物土壤某些种类的微生物数量多但种群丰富度低,属于土壤微生物利用碳源的理论模式III。由此可见,在“低”(406 m、550 m和591m)、“中”(745 m)、“高”(800 m、1025 m和1355 m)海拔处呈现了不同的碳源利用模式,据此推测鹦哥岭热带山地雨林不同海拔处土壤微生物群落结构不同。

Figure 1. AWCD dynamics change with incubation time at different altitudes
图1. 不同海拔下的土壤微生物群落平均颜色变化率随时间的变化
3.3. 不同海拔土壤微生物利用不同碳源的强度特征
不同海拔热带雨林样地土壤微生物群落对碳水化合物类、羧酸类、氨基酸类、多聚物类、酚酸类和胺类六大类碳源的利用结果表明(图2-A,图2-B),鹦哥岭热带山地雨林各海拔点样地土壤微生物群落对碳源的利用情况不同,不同海拔样地土壤微生物群落对碳源的利用程度不同,且南坡和北坡存在差异。海拔和坡向共同影响土壤微生物利用碳源的强度。
从图2-A可知,南坡不同海拔土壤微生物群落利用胺类和羧酸类碳源差异不显著(p > 0.05),对胺类碳源的利用在550 m海拔处最高,对羧酸类碳源的利用在1025 m海拔处最高。550 m、745 m、1025 m和1355 m四个海拔样地土壤微生物群落对多聚物类碳源的利用差异不显著(p > 0.05),但均显著高于406 m海拔处(p < 0.05),且在1355 m海拔处最高。土壤微生物群落对碳水化合物、氨基酸类、酚酸类碳源的利用随海拔的升高先升高后降低,1025 m海拔处的样地土壤微生物群落对酚酸类碳源的利用最高,1025 m和1355 m两个“高”海拔处的样地土壤微生物群落对酚酸类碳源的利用显著高于406 m的“低”海拔处(p < 0.05),745 m的“中”海拔处样地土壤微生物群落对酚酸类碳源的利用与406 m和550 m 两个“低”海拔处差异不显著(p > 0.05)。550 m海拔处土壤微生物群落对碳水化合物类和氨基酸类碳源的利用最高,其中:对碳水化合物类碳源利用,550 m海拔处显著高于1355 m海拔处(p < 0.05),与406 m、745 m和1025 m海拔处差异不显著(p > 0.05);对氨基酸类碳源的利用,550 m海拔处显著高于406 m海拔处和1355 m海拔处(p < 0.05),与745 m和1025 m海拔处差异不显著(p > 0.05)。南坡不同海拔土壤微生物群落利用不同碳源(胺类和羧酸类除外)的不同,同样也说明了南坡不同海拔土壤微生物群落结构不同。
从图2-B可知,北坡不同海拔土壤微生物群落对碳水化合物、氨基酸类、多聚物类、酚酸类、胺类和羧酸类六大类碳源的利用均随海拔的升高先升高后降低,其中:利用氨基酸类、酚酸类和胺类碳源差异不显著(p > 0.05),且在591 m海拔处最高;对碳水化合物类碳源的利用,591 m的“中”海拔处显著高于458 m的“低”海拔和800 m的“高”海拔处(p < 0.05),“低”海拔处和“高”海拔处差异不显著(p > 0.05);对多聚物类碳源的利用,591 m的“中”海拔处显著高于800 m的“高”海拔处(p < 0.05),“低”海拔处和“高”海拔处差异不显著(p > 0.05);对羧酸类碳源的利用,591 m的“中”海拔处显著高于458 m的“低”海拔处(p < 0.05),“低”海拔处和“高”海拔处差异不显著(p > 0.05)。北坡不同海拔处土壤微生物群落利用不同碳源(氨基酸类、酚酸类和胺类除外)的不同同样也说明了北坡不同海拔土壤微生物群落结构不同。
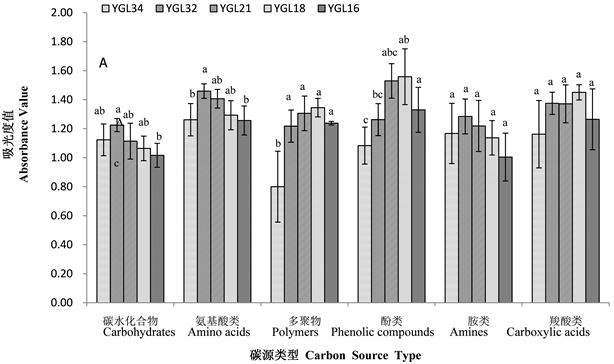
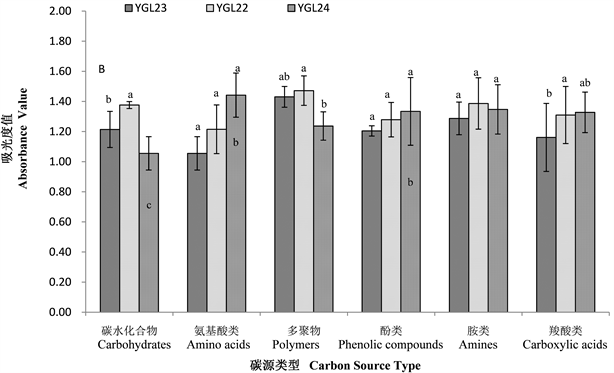
Figure 2. Different carbon source utilization by soil microbial community in different slope directions (A: Southern slope, B: northern slope) at different altitudes
图2. 不同坡向(A:南坡,B:北坡)不同海拔土壤微生物群落对不同碳源的利用
3.4. 不同海拔土壤微生物群落功能多样性指数特征
从表5可知,鹦哥岭热带山地雨林土壤微生物群落McIntosh指数和Shannon指数整体随海拔的升高先升高后降低,McIntosh指数在591 m海拔处显著最高,部分海拔样地间差异显著(p < 0.05);Shannon指数在591 m海拔处最高,各海拔样地间差异均不显著(p > 0.05);Simpson指数和Pielou指数在各海拔样地间差异均不显著(p > 0.05)。结合图1,可知591 m海拔处(YGL22)热带山地雨林土壤微生物活性最强。
Table 5. Functional diversity index of soil microbial communities at different altitudes
表5. 不同海拔的土壤微生物群落功能多样性指数
注:表中同一列字母不同,表示差异显著(p < 0.05)。
3.5. 不同海拔土壤微生物群落代谢功能主成分分析
提取Biolog-ECO微平板上144 h的31种碳源底物利用情况进行主成分分析,结果表明,不同海拔土壤微生物第一主成分(PC1)和第2主成分(PC2)贡献率分别为32.38%和28.47%,累计贡献率达60.75%,是微生物群落碳源利用变异的主要来源,其他主成分存在一定的贡献率(图3)。YGL34位于PC1的负端得分系数分别为−1.9228;YGL32、YGL21、YGL22和YGL23均位于位于PC1轴“基点”附近,得分系数在−0.2119~0.94139之间,四样点基本聚集为一簇;YGL18、YGL16、YGL24均位于PC2的正端,得分系数在0.6108~1.3135之间,三样点基本聚集为一簇。另外,YGL34位于PC2的正端,得分系数为0.3245,为单独的一簇。由此可见,PC1和PC2基本能区分鹦哥岭不同海拔土壤微生物的群落特征,“低”(406 m)、“中”(458 m, 550 m, 591 m, 745 m)、“高”(800 m, 1025 m, 1355 m)海拔显著影响土壤微生物的碳源利用。
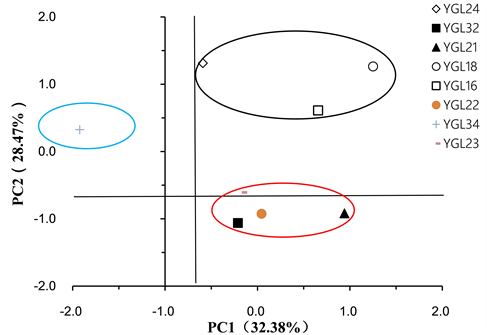
Figure 3. Principle component (PC) analysis for carbon utilization of soil microbial communities at different altitudes
图3. 不同海拔土壤微生物群落碳源利用主成分分析
进一步将主成分得分系数与单一碳源AWCD值做相关分析,即获得31种碳源在2个主成分上的载荷值,载荷因子越高,表示该碳源对主成分的影响越大,两者之间的相关性越高。从表6可知,鹦哥岭不同海拔热带山地雨林土壤微生物,在31种碳源中与PC1高度相关(主成分相关系数 > 0.7)的碳源有7种,其中5种正相关,2种负相关,分别属于碳水化合物类3种,多聚物类2种,羧酸类和酚酸类各1种;与PC2高度相关的碳源有8种,其中5种正相关,3种负相关,分别属于碳水化合物类4种,氨基酸酸类2种,羧酸类和多聚物类各1种。综合与PC1和PC2高度相关的碳源类型可见,在主成分分离中起主要贡献作用的是碳水化合物类、多聚物类和羧酸类的碳源。
Table 6. Loading factors of carbon sources utilization of soil microbial in tropical mountain rain forest of Yinggeling
表6. 鹦哥岭热带山地雨林土壤微生物主要利用碳源的载荷因子
3.6. 不同海拔土壤微生物群落碳代谢与土壤理化性质的相关性分析
从表7可知,鹦哥岭热带山地雨林土壤微生物群落功能多样性McIntosh指数与土壤理化性质相关性程度大小依次为过氧化氢酶 > 有效磷 > 酸性磷酸酶 > 有机质 > 全氮 > 铵态氮 > 全磷 > 脲酶> pH > 硝态氮 > 速效钾 > 碱解氮 > 全钾。其中:与pH、全钾、有效磷、速效钾正相关,与其他理化因子之间负相关;与有效磷显著正相关(p < 0.05),与过氧化氢酶、酸性磷酸酶显著负相关(p < 0.05)。土壤微生物群落功能多样性Shannon指数、Simpson指数和Pielou指数与土壤理化性质相关性绝对值均较小(r < 0.2),极弱相关或不相关。
Table 7. Correlation analysis of physical-chemical properties and microbial community functional diversity in the tropical montane rainforest of Yinggeling
表7. 鹦哥岭热带山地雨林土壤理化及酶活性与微生物群落功能多样性的相关性
注:n = 24,*表示显著相关(p < 0.05),**表示极显著相关(p < 0.01)。
3.7. 不同海拔土壤微生物群落碳代谢与凋落物理化性质的相关性分析
Table 8. Correlation between physical—chemical properties of litter and functional diversity of microbial community
表8. 凋落物理化性质与微生物群落功能多样性的相关性
注:n = 24,*表示显著相关(p < 0.05)。
凋落物是联系植被与土壤两个系统的纽带,凋落物分解是森林生态系统尤为关键的生态过程之一,对森林的生物地球化学循环起着十分重要的作用,是土壤有机质的主要来源。凋落物的化学组成影响土壤微生物学性状,为了解凋落物与土壤微生物群落功能多样性之间的关系,对土壤微生物群落功能多样性与凋落物理化性质进行相关性分析。从表8可以看出,鹦哥岭热带山地雨林土壤微生物群落功能多样性McIntosh指数与凋落物有机碳含量显著负相关(p < 0.05),与凋落物其他理化性质之间弱相关或极弱相关,甚至不相关;土壤微生物群落功能多样性Shannon指数、Pielou指数和Simpson指数与凋落物理化性质弱相关或极弱相关,甚至不相关。
4. 讨论
4.1. 海拔变化对土壤微生物利用碳源的影响
海拔梯度的上升会引起气候、植被和土壤养分等环境因子的变化,尤其是引起直接作用于生境的光照、湿度、温度等气候生态因子的变化,最终影响土壤微生物群落活性和多样性特征。本研究中不同海拔梯度下土壤微生物群落的AWCD值均随时间的延长而上升,整体上随海拔上升呈现先上升后下降的单峰变化趋势,该研究结果与王颖、朱平、孟苗婧等的研究结果基本一致 [10] [34] [35] ,与吴则焰、韩冬雪、宋贤冲的研究结果不一致 [36] [37] [38] [39] 。由此推测,本研究中鹦哥岭热带山地雨林不同海拔土壤微生物群落的AWCD值的变化特征可能是土壤理化性质和土壤酶活性综合作用的结果,土壤有效磷、过氧化氢酶和酸性磷酸酶活性是最重要的影响因子。
4.2. 海拔变化对土壤微生物群落功能多样性指数的影响
土壤中微生物物种数量及其分布的均匀度决定土壤微生物群落中的物种多样性,物种多样性指数的差异侧面反映了土壤微生物多样性,群落多样性指数可用来指示土壤微生物群落利用碳源的程度 [40] 。山地生态学中的“中部膨胀”理论认为植物和动物在垂直地带上的多样性分布具有明显的中峰优势,这一现象已被以往的研究证实,微生物在垂直地带上的多样性分布同样存在类似的中锋优势分布规律 [36] [41] 。本研究中鹦哥岭热带山地雨林生态系统的土壤微生物群落功能多样性指数整体随海拔的升高先增加后减少,土壤微生物群落功能多样性在垂直地带上的分布具有明显的中锋优势;同时发现Shannon、Simpson和Pielou指数在各海拔间差异均不显著,McIntosh指数仅在部分海拔间差异显著;591 m海拔处样地的Shannon指数和McIntosh指数均最高,406 m海拔处样地的Shannon指数最低。由此可见,鹦哥岭热带山地雨林“中”海拔处样地微生物种类多且分布均匀,利用碳源多且强度大;鹦哥岭热带山地雨林“低”海拔处样地微生物种类相对较少且分布不均匀。结合土壤及凋落物理化性质及酶活性与微生物群落功能多样性的相关性结果(表7~8),推测鹦哥岭热带山地雨林土壤微生物群落功能多样性的垂直地带性差异可能与植被类型的改变及其伴随的土壤养分的改变有关。
4.3. 海拔变化对土壤微生物群落利用碳源强度和利用模式的影响
海拔高度变化影响植被组成、结构,土壤温湿度、水分、养分,土壤有机质和微生物活性,进而影响土壤微生物群落功能和结构 [42] [43] [44] [45] 。本研究发现鹦哥岭热带山地雨林南坡的“低”海拔处,土壤微生物群落对氨基酸类和羧酸类碳源的利用程度相对较高,对多聚物类碳源的利用程度相对较低;南坡的“中、高”海拔处,土壤微生物群落对酚类和羧酸类碳源的利用程度相对较高,对碳水化合物类的利用程度相对较低。北坡各海拔处的土壤微生物群落对氨基酸类碳源的利用程度相对较高,对碳水化合物类碳源的利用程度相对较低。上述研究结果与宋贤冲、吴则焰、金裕华等的研究结果不同 [38] [39] [40] 。土壤输入性碳源的种类影响土壤微生物群落对碳源的利用强度和利用模式 [46] 。本研究中南坡745 m海拔处样地土壤微生物群落对碳源的利用模式为理论模式I,南坡1025 m、1355 m与北坡458 m、800 m海拔处样地土壤微生物群落对碳源的利用模式为理论模式II,南坡406 m、550 m和北坡591 m海拔处样地土壤微生物群落对碳源的利用模式为理论模式III。植物群落组成和结构可能通过影响其根际土壤的理化性质,进而影响土壤微生物群落组成和结构,最终导致不同海拔处土壤微生物功能多样性的差异化 [47] 。结合不同海拔样地植被及土壤理化性质特点,推测在诸多环境因子的多重及交互作用的复杂关系中,植被和土壤要素对不同海拔热带山地雨林土壤微生物的碳源利用模式和强度产生重要的影响。
4.4. 海拔变化对土壤微生物群落碳源代谢多样性的影响
主成分分析可以直观的展示土壤微生物群落功能的相似状况和综合差异,碳源底物的初始载荷因子体现了主成分与碳源利用的相关性,初始载荷因子的高低反映了该碳源对主成分的影响程度 [35] 。本研究发现海拔梯度显著影响鹦哥岭热带山地雨林土壤微生物的碳源利用,土壤微生物群落碳源代谢多样性的垂直地带性差异主要体现在对碳水化合物类、多聚物类和羧酸类碳源的利用,该结果与以往大多数的研究结果不一致 [36] [37] [38] 。众所周知,碳水化合物类的碳源既是自然界矿化过程的主要物质,也为大多数异养微生物提供有机营养物;植物根系分泌物羧酸和小分子的氨基酸等碳源类物质 [48] 。鹦哥岭热带山地雨林特殊生境提供的高温、高湿环境为土壤微生物的生命活动提供了良好的条件,可能有利于土壤微生物对多聚物类等难氧化碳源的利用。据此推测,鹦哥岭热带山地雨林土壤微生物碳源代谢多样性的垂直地带性差异可能与热带山地雨林高温、高湿的特殊生境以及土壤养分的改变有关。
4.5. 环境因子对土壤微生物群落功能多样性的影响
气候条件、土壤理化性质及植物群落等环境因子影响土壤微生物群落结构和组成,由于气候、土壤和植物因子的多重或交互作用导致土壤微生物多样性呈现相应的时空变化。众多的研究发现有机碳、pH、全氮、全磷和有效磷等土壤理化性质的变化影响土壤微生物功能多样性的空间变化 [10] [35] 。本研究中土壤微生物群落功能多样McIntosh指数与有效磷显著正相关,与过氧化氢酶和酸性磷酸酶活性显著负相关;土壤微生物群落功能多样性Shannon指数、Simpson指数和Pielou指数(E)与土壤理化性质极弱相关,甚至不相关。据此推测,土壤有效磷、过氧化氢酶和酸性磷酸酶活性是鹦哥岭热带山地雨林土壤微生物群落功能多样性随海拔梯度变化最主要的影响因子。
4.6. 凋落物对土壤微生物群落功能多样性的影响
凋落物是热带山地雨林森林生态系统生产力构成中的重要组成成分,是热带山地雨林生态系统物质和能量的主要来源,也是土壤与微生物间的物质循环枢纽 [49] 。土壤微生物以分解者角色,积极参与森林生态系统中的物质循环和能量流动,是凋落物分解、土壤呼吸以及生物化学循环等过程的主要驱动因子之一 [50] 。本研究发现鹦哥岭热带山地雨林土壤微生物群落功能多样性McIntosh指数与凋落物有机碳含量显著负相关,与凋落物其他指标相关性极弱;Shannon指数、Pielou指数和Simpson指数与凋落物各项理化指标极弱相关。据此推测鹦哥岭热带山地雨林生态系统中凋落物的输入或去除显著影响土壤微生物群落功能多样性的McIntosh指数,但对土壤微生物群落的Shannon多样性指数、Pielou均匀度指数和Simpson指数影响不大,该结果与以往的研究结果存在一定的相似性 [47] 。另外,本研究还发现各海拔样地凋落物干重差异不显著,但是凋落物的有机碳含量仅在部分海拔间差异显著,据此推测凋落物对热带山地雨林土壤微生物群落碳代谢多样性的影响在于凋落物碳的可利用性,即凋落物碳的质量,尤其是有机碳是影响土壤微生物群落功能多样性的重要环境因子。
5. 结论
鹦哥岭热带山地雨林土壤和凋落物理化性质、土壤酶活性的海拔分布具有异质性,部分海拔样地间差异显著,且南坡和北坡不同。不同海拔样地土壤微生物群落的AWCD值不同,pH是重要的影响因子。不同海拔样地土壤微生物对碳源的利用模式和利用程度不同,土壤微生物碳源代谢多样性的垂直地带性差异主要体现在碳水化合物、多聚物类和羧酸类碳源的利用上,且在部分海拔间差异显著。土壤微生物群落功能多样性指数随海拔的升高先增加后减少,符合“中峰分布”格局。土壤微生物群落功能McIntosh指数在部分海拔间差异显著,与土壤有效磷含量显著正相关,与土壤过氧化氢酶、酸性磷酸酶、凋落物有机碳含量显著负相关,且南坡和北坡不同;各海拔样地土壤微生物群落功能Shannon指数、Simpson指数和Pielou指数差异不显著,与土壤和凋落物理化性质以及土壤酶活性弱相关,甚至不相关。土壤有效磷、酸性磷酸酶、过氧化氢酶和凋落物有机碳含量是导致鹦哥岭热带山地雨林土壤微生物群落功能多样性的海拔分布异质性的重要影响因子。
基金项目
海南省自然科学基金面上项目(320MS040):鹦哥岭热带雨林土壤微生物多样性及其影响因子的研究。
NOTES
*通讯作者。