摘要: 研究以武汉市八分山、青龙山、马鞍山、磨山和九峰山森林公园为对象,研究土壤微生物量碳的分布特征及其土壤环境影响因子。在每座山选择8种主要树种,共设置120个采样点,于2021年9月至11月进行土壤样品采集,采用抖落法采集根际土壤样品。结果显示:(1) 八分山(57.42~502.22 mg∙kg−1)、青龙山(148.59~563.78 mg∙kg−1)、马鞍山(270.76~908.44 mg∙kg−1)和磨山(137.45 ~ 654.81 mg∙kg−1)不同植物群落土壤微生物量碳间差异显著,九峰山(224.18~595.56 mg∙kg−1)不同植物土壤微生物量碳间无显著差异。(2) 青龙山土壤微生物量碳与铵态氮(r = 0.462, p < 0.05)呈显著正相关;马鞍山土壤微生物量碳与有机碳(r = 0.648, p < 0.01)、硝态氮(r = 0.434, p < 0.05)、铵态氮(r = 0.474, p < 0.05)、无机氮(r = 0.532, p < 0.01)、C:P (r = 0.560, p < 0.01)和N:P (r = 0.420, p < 0.05)呈显著正相关;九峰山土壤微生物量碳与铵态氮(r = 0.578, p < 0.01)、无机氮(r = 0.573, p < 0.01)、C:P (r = 0.551, p < 0.01)和N:P (r = 0.465, p < 0.05)呈显著正相关;八分山和磨山土壤微生物量碳均与环境因子无显著差异。(3) 五座森林公园的土壤微生物量熵碳范围在1%~4%。层次分割法表明土壤微生物量碳的关键驱动因子为有机碳。
Abstract:
To analyze the distribution characteristics and influencing factors of soil microbial carbon, and to explore the relationship between soil microbes and environmental factors. The study was con-ducted in the forest parks of Bafen Mountain, Qinglong Mountain, Maanshan Mountain, Mashan Mountain and Jiufeng Mountain in Wuhan City, and eight major constituent tree species were se-lected for each mountain, with a total of 120 sampling points, and soil samples were collected from September to November 2021, and inter-root soil samples were collected by the shaking drop method. The results showed that: (1) There were no significant differences in soil microbiomass carbon among different plant communities in Bafen Mountain 57.42~502.22 mg∙kg−1), Qinglong Mountain(148.59~563.78 mg∙kg−1), Ma’anshan Mountain (270.76~908.44 mg∙kg−1), and Mashan Mountain (137.45 ~ 654.81 mg∙kg−1) showed significant differences among soil microbial mass carbon of different plant communities, and Jiu Feng Shan (224.18~595.56 mg∙kg−1) showed non-significant differences among soil microbial mass carbon of different plants. (2) There was a significant positive correlation between soil microbial and ammonium nitrogen (r = 0.462, p < 0.05) in Qinglong, and a significant positive correlation between soil microbial amount carbon and organic carbon (r = 0.648, p < 0.01), nitrate nitrogen (r = 0.434, p < 0.05), ammonium nitrogen (r = 0.474, p < 0.05), inorganic nitrogen (r = 0.532, p < 0.01), C:P (r = 0.560, p < 0.01) and N:P (r = 0.420, p < 0.05) were significantly positively correlated; soil microbial amount carbon in Jiufeng was significantly positively correlated with ammonium nitrogen (r = 0.578, p < 0.01), inorganic nitrogen (r = 0.573, p < 0.01), C:P (r = 0.551, p < 0.01) and N:P (r = 0.465, p < 0.05) were significantly positively correlated; neither Bafen nor Moshan soil microbiomass carbon was significantly related to environmental factors. (3) Soil microbiomass entropy carbon in the five forest parks ranged from 1% to 4%. Hierarchical partitioning was further used to derive that the driver of soil microbiomass carbon was organic carbon.
1. 引言
森林生态系统具有巨大的固碳潜力。森林植被存储着全球植被总碳储量的86%,森林土壤存储着全球土壤总储存量的73% [1] 。土壤微生物量碳(Microbial biomass carbon, MBC)是土壤微生物生物量主要的组成,不仅对土壤有机质和养分有重要作用,还是一个直接调控土壤养分影响植被生长的重要活性养分库,因此,土壤MBC是土壤肥力变化的重要指标之一 [2] [3] 。研究表明,不同树种凋落量及根系分泌物 [4] 、土壤深度 [5] 、季节变化 [6] 等会影响土壤MBC的差异。城市森林是城市生态系统的重要组成部分,在城市可持续发展中发挥巨大的生态功能,如净化空气,改善城市“热岛”问题等 [7] ,也是区域碳库的关键组成。
武汉市森林面积179.2万亩,森林蓄积量840.87 m3 [8] 。但是武汉市森林公园土壤微生物量碳特征研究较为缺乏,多数研究以湖北省主要森林类型的土壤理化性质及质量为主 [9] [10] 。因此,还需研究森林土壤MBC与土壤碳库之间的关系,土壤与植物之间的机制,探讨城市森林的生态作用,研究土壤碳汇潜力。论文以武汉市的八分山、青龙山、马鞍山、磨山和九峰山森林公园为对象,探究森林公园不同植物根际土壤微生物生物量碳的分布特征及其影响因素,为了解城市森林公园土壤碳汇变化规律,揭示土壤碳汇形成机制提供科学依据。
2. 研究区概况
武汉市(113˚41'~115˚05'E, 29˚58'~31˚22'N)位于湖北省,属亚热带季风性湿润气候区,日照充足,热量丰富,雨热同季,降雨集中等特点。年降雨量为1205 hm,活动积温在5000~5300℃,年无霜期达240天。植物区系属中亚热带常绿阔叶林与落叶阔叶林混交林。土壤以红黄壤、棕壤、黄褐土为主,母岩多为砂岩,存在零星石灰岩。八分山森林公园(114˚01'~114˚35'E, 29˚58'~30˚32'N)位于武汉江夏区纸坊街,海拔272.3 m,森林覆盖率高达96%;青龙山森林公园(113˚50'~114˚22'E, 30˚33'~30˚17'N)位于江夏区纸坊街,森林覆盖率高达95%以上;马鞍山森林公园(114˚25'~114˚27'E, 30˚30'~30˚32'N)位于武汉东湖吹笛风景区,森林覆盖率达80%;磨山森林公园(114˚40'~114˚43'E, 30˚54'~30˚56'N)位于东湖磨山西峰景区;九峰森林公园(114˚29'~114˚30'E, 30˚30'~30˚31'N)位于武汉市洪山区,海拔高度51.2~202 m,森林覆盖率85%以上。
3. 研究方法
3.1. 样品采集
于2021年9~11月在八分山、青龙山、马鞍山、磨山和九峰山森林公园选取代表性强、植被类型相对单一以及人为干扰相对较小的森林样带,每座山设置24个样点,共120个样点(图1)。剔除石块、根系、凋落物、动植物残体等,过2 mm土壤筛后将样品一分为二,一部分低温保存带回实验室,置于−20℃冰箱保存,用于测定土壤微生物生物量碳含量;另一部分自然风干、粉碎、过筛(0.25 mm)后,用于测定土壤理化性质。
3.2. 土壤指标测定
土壤MBC采用氯仿熏蒸–硫酸钾浸提法测定 [11] [12] [13] ,将经过氯仿熏蒸和未熏蒸的新鲜土壤用0.5 mol∙L−1的K2SO4溶液浸提,再用TOC仪(耶拿multi N/C 2100,德国)测定。土壤理化性质按土壤农化分析与环境检测中的方法进行测定 [14] 。速效磷(AP)采用钼锑抗比色法测定;有机碳(SOC)采用H2SO4-K2Cr2O7外加热法测定;铵态氮(
-N)与硝态氮(
-N)采用流动分析仪法(Skalar San++,荷兰);pH值采用1:2.5土水比电位法;土壤微生物量熵碳(qMBC)为土壤MBC与SOC比 [15] 。
3.3. 数据分析方法
利用IBM SPSS Statistics 26软件对不同样地的土壤MBC、去、qMBC、SOC、AP、
-N、
-N、SIN等土壤指标进行方差分析(One-Way ANVON)检验土壤指标间的差异显著性,平均数比较采用最小显著数测验法,显著水平均为0.05。皮尔森相关性分析确定土壤MBC与SOC、AP、
-N、
-N、SIN、C:P、C:N、N:P间的相关性。为克服确定权重的主观性,提供指标赋权的一致性,通过R语言包(rdacca.hp)分析不同植物根际土壤MBC的驱动因子。用Origin 2021软件制图。
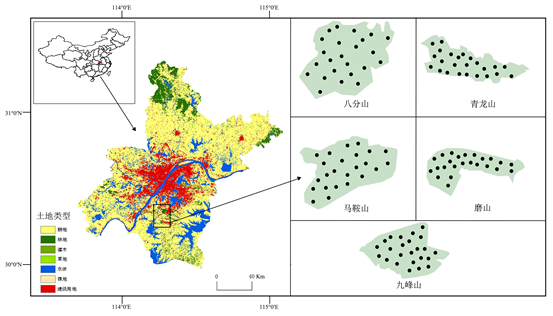
Figure 1. Sampling map of the Wuhan forest parks
图1. 武汉市森林公园采样图
4. 结果与分析
4.1. 土壤微生物生物量碳含量特征
不同植物群落土壤MBC存在显著性差异(图2)。八分山土壤MBC为57.42 ± 16.29 (锥栗)~502.22 ± 57.10 mg∙kg−1(构树、马尾松)。青龙山土壤MBC为148.59 ± 52.65 (檵木)~ 563.78 ± 28.53 mg∙kg−1 (锥栗)。马鞍山土壤MBC为270.76 ± 77.03 (桂花)~908.44 ± 163.16 mg∙kg−1 (香樟),群落土壤MBC含量最高。磨山土壤MBC为137.45 ± 61.03 (香樟)~654.81 ± 89.83 mg∙kg−1 (锥栗)。九峰山土壤MBC为224.18 ± 74.84 (杉松)~595.56 ± 131.51 mg∙kg−1 (黑足鳞毛蕨),不同植物群落之间差异不显著。
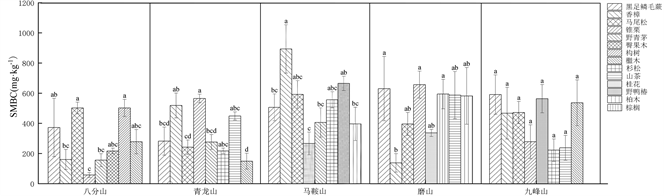 注:不同小写字母代表差异显著(p < 0.05)。
注:不同小写字母代表差异显著(p < 0.05)。
Figure 2. Carbon content of microbial biomass in rhizosphere soil in the Wuhan forest parks
图2. 武汉市森林公园植物根际土壤微生物量碳含量
4.2. 土壤养分含量与化学计量比特征
不同植物根际土壤理化性质间均存在显著差异(表1)。八分山、青龙山、马鞍山、磨山和九峰山根际土壤有机碳(SOC)含量范围分别为18.88 ± 0.55 (构树)~36.14 ± 3.69g∙kg−1(野青茅)、10.47 ± 2.23 (香樟)~27.58 ± 2.74 g∙kg−1 (锥栗)、18.77 ± 1.81 (黑足鳞毛蕨)~46.02 ± 4.69 g∙kg−1 (香樟)、17.31 ± 1.55 (马尾松)~29.40 ± 2.36 g∙kg−1 (香樟)和11.74 ± 2.16 (山茶)~31.13 ± 2.56 g∙kg−1(野鸭椿);速效磷(AP)分别为10.4 ± 1.88(野青茅)~46.59 ± 3.19 g∙kg−1 (构树)、73.73 ± 13.11 (杉松)~32.18 ± 5.28 g∙kg−1 (檵木)、31.82 ± 3.87 (柏木)~96.90 ± 4.24 g∙kg−1 (桂花)、25.40 ± 1.82 (黑足鳞毛蕨)~99.90 ± 1.84 g∙kg−1 (桂花)和19.97 ± 0.31 (檵木)~106.54 ± 17.53 g∙kg−1 (杉松);铵态氮(
-N)分别为6.27 ± 0.99 (马尾松)~14.53 ± 3.73 g∙kg−1 (野青茅)、5.17 ± 0.39 (檵木)~8.53 ± 1.44 g∙kg−1 (黑足鳞毛蕨)、7.20 ± 0.41 (马尾松)~15.33 ± 1.77 g∙kg−1 (香樟)、4.10 ± 1.02 (马尾松)~9.33 ± 1.26 g∙kg−1 (桂花)和3.67 ± 0.14 (山茶)~9.73 ± 0.75 g∙kg−1 (马尾松);硝态氮(
-N)分别为2.93 ± 1.63 (香樟)~8.73 ± 3.98 g∙kg−1 (构树)、1.60 ± 0.61(香樟)~11.65 ± 0.35 g∙kg−1 (锥栗)、1.07 ± 0.39 (桂花)~9.17 ± 3.31 g∙kg−1 (野青茅)、3.17 ± 0.96 (山茶)~16.80 ± 5.77 g∙kg−1 (锥栗)和3.47 ± 1.26 (马尾松)~6.83 ± 1.46 g∙kg−1 (檵木);无机氮(SIN)分别为12.07 ± 1.12 (臀果木)~22.43 ± 7.05 g∙kg−1 (野青茅)、7.07 ± 0.72 (香樟)~18.85 ± 0.35 g∙kg−1 (锥栗)、10.77 ± 1.03 (桂花)~24.33 ± 1.70 g∙kg−1 (香樟)、9.73 ± 1.20 (山茶)~22.47 ± 5.44 g∙kg−1 (锥栗)和8.27 ± 0.71 (锥栗)~14.10 ± 2.26 g∙kg−1 (檵木)。
土壤pH范围介于4.19~5.74之间,五座森林公园植物根际土壤为酸性。

Table 1. Physical and chemical indexes of plant rhizosphere soil in Wuhan forest parks
表1. 武汉市森林公园植物根际土壤理化指标
注:不同小写字母代表差异显著(p < 0.05)。
不同植物根际的土壤养分的化学计量比均存在显著差异(表2)。八分山、青龙山、马鞍山、磨山和九峰山根际土壤C:P范围分别为412.30 ± 35.36 (构树)~4400.52 ± 2619.53 (黑足鳞毛蕨)、155.74 ± 46.38 (香樟)~686.67 ± 196.25 (檵木)、206.89 ± 21.89(桂花)~1320.75 ± 180.49 (香樟)、250.43 ± 48.25 (桂花)~1131.57 ± 113.97 (黑足鳞毛蕨)和236.93 ± 103.39 (杉松)~1093.53 ± 217.99 (檵木);C:N分别为1105.61 ± 371.85 (构树)~2325.96 ± 70.94 (臀果木)、1219.60 ± 117.58 (檵木)~1982.62 ± 302.33 (马尾松)、1112.92 ± 50.02 (杉松)~1998.98 ± 88.47 (野鸭椿)、1143.16 ± 222.52 (柏木)~2300.88 ± 414.27 (山茶)和1157.31 ± 173.45 (山茶)~3497.00 ± 996.02 (野鸭椿);N:P分别为0.30 ± 0.04 (臀果木)~2.95 ± 1.82 (黑足鳞毛蕨)、0.10 ± 0.02 (香樟)~0.57 ± 0.17 (檵木)、0.11 ± 0.01(桂花)~0.70 ± 0.07 (香樟)、0.14 ± 0.01(桂花)~0.88 ± 0.13 (黑足鳞毛蕨)和0.10 ± 0.02 (野鸭椿)~0.71 ± 0.12(檵木)。
Table 2. Stoichiometric ratio of soil nutrients in rhizosphere of plants in the Wuhan forest parks
表2. 武汉市森林公园植物根际土壤养分含量的化学计量比
注:不同小写字母代表差异显著(p < 0.05)。
4.3. 土壤微生物生物量熵碳变化特征
不同植物根际土壤微生物生物量熵碳(qMBC)的变化范围介于1%~4% (图3)。其中青龙山香樟群落的qMBC值最高,八分山锥栗群落qMBC值最低。
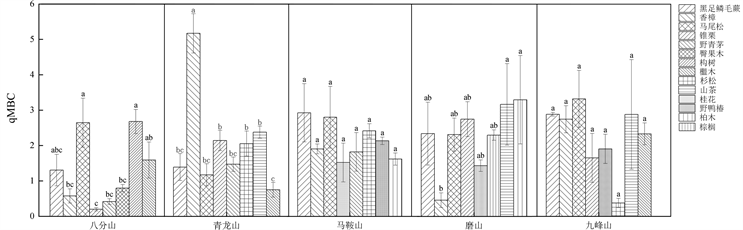 注:不同小写字母代表差异显著(p < 0.05)。
注:不同小写字母代表差异显著(p < 0.05)。
Figure 3. Entropy carbon of plant rhizosphere soil microbial biomass in Wuhan forest park
图3. 武汉市森林公园植物根际土壤微生物生物量熵碳
4.4. 土壤MBC与土壤环境的相关性
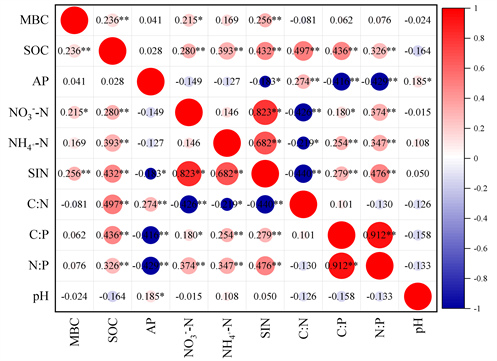
土壤MBC与土壤环境之间的相关关系见图4。土壤MBC与SOC、土壤无机氮(SIN)呈极显著正相关,与
−N呈显著正相关;SOC与
−N、
−H、SIN、C:N、C:P、N:P呈极显著正相关。
 *.在0.05级别(双尾),相关性显著。**.在0.01级别(双尾),相关性显著。
*.在0.05级别(双尾),相关性显著。**.在0.01级别(双尾),相关性显著。
Figure 4. Correlation heat map of soil factors in plant rhizosphere in the Wuhan forest park
图4. 武汉市森林公园植物根际土壤因子的相关性热图
4.5. 土壤理化性质对土壤MBC的单一变量解释率
采用层次分割法(rdacca.hp)分析土壤MBC的各因子解释率(图5),只有SOC的p值小于0.05,所以土壤MBC的关键驱动因子是SOC。土壤因子AP、C:P、N:P和pH贡献程度较低,解释率为负值。
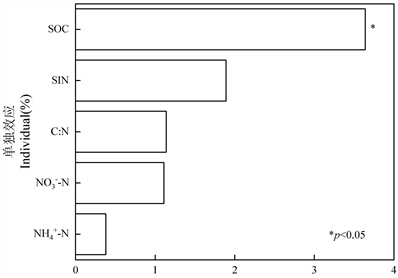
Figure 5. Individual explanatory rate of soil factors
图5. 土壤因子的单一解释率
5. 讨论
5.1. 土壤MBC的分布特征
植物通过凋落物和根际分泌物向土壤微生物提供营养,土壤微生物通过转化有机物供植物利用 [16] 。因此,和贾国梅等 [17] 研究类似,不同植物群落间根际土壤MBC间具有显著差异。本研究中森林公园土壤MBC均低于全球土壤MBC水平(347~971 mg∙kg−1) [18] 。我们进一步发现乔木类植物群落的根际土壤MBC最高,例如八分山马尾松(乔木)和锥栗(大乔木)、青龙山锥栗(大乔木)、马鞍山香樟(大乔木)、磨山锥栗(大乔木)群落MBC含量最高(图2)。
土壤qMBC是土壤微生物量碳对土壤营养库的贡献率 [19] ,表征有机碳积累或损失,土壤MBC与SOC的比值越大说明土壤微生物固碳能力越强,有机碳积累越多 [20] 。本研究发现:森林公园qMBC值在1%~4%,与平均土壤qMBC (1%~3%)相符 [16] ,大的变化范围表明研究区域具有较高的固碳潜力。不同森林公园中不同树种的qMBC值表明(图3)草本、乔木和灌木类植物群落处于有机碳积累阶段,固碳潜力较强。植被类型不同,植被表层土壤的凋落物及植被根系分泌物存在差异,土壤微生物的结构和多样性存在差异。
5.2. 土壤MBC与土壤因子之间的相关关系
已有研究表明微生物残体是SOC的重要来源,微生物残体对SOC的平均贡献约为35%,并且微生物途径对SOC的积累与土层的深度呈正相关,但在森林表层土壤(0~20 cm)中,SOC积累的主导因素为植物途径 [21] 。周莎等 [22] 研究发现不同纬度森林土壤MBC与SOC呈极显著正相关,李万年等 [20] 研究证实SOC是影响土壤MBC的主要因子,这与本文研究结果一致。土壤无机氮(
-N和
-N)是植物吸收利用的主要氮素。赵高山等 [23] 研究发现土壤MBC与碱解氮呈极显著正相关,与本文结果一致。张雅茜等 [24] 对亚热带丘陵区大山冲林场研究发现土壤MBC与AP呈极显著正相关,这与本文研究结果不一致,这可能是因为土壤MBC与AP之间关系受许多外在因素的影响,比如土壤pH、植物量等。土壤pH均与土壤MBC无显著相关关系,这可能与土壤类型和土壤微生物活性差异等因素有关 [25] 。
土壤C:P、C:N、N:P化学计量比能够表征植物生长及土壤养分利用情况。本研究区域的C:P和C:N值均高于我国土壤C:P(12)和C:N(61) [26] ,说明土壤有机质积累速率高于分解速率,养分净固持能力加强。N:P值低于中国土壤N:P平均值9.3 [26] ,说明研究区域土壤氮限制严重 [27] 。
6. 结论
(1) 八分山、青龙山、马鞍山、磨山和九峰山森林公园土壤MBC分别为57.42~502.22 mg∙kg−1、148.59~563.78 mg∙kg−1、270.76~908.44 mg∙kg−1、137.45~654.81 mg∙kg−1、224.18~595.56 mg∙kg−1。
(2) 土壤MBC与SOC、土壤无机氮(SIN)呈极显著正相关,与
−N呈显著正相关。其中SOC是影响土壤MBC的主要驱动因子。
(3) 土壤qMBC值为1%~4%,研究区固碳潜力较高。
研究加入土壤MBC与碳汇之间的联系,能更加全面的对亚热带城市森林土壤碳汇分布及潜力进行分析。后续研究需要进一步加强高固碳能力树种的固碳驱动因素及固碳潜力核算,为区域森林生态系统碳增汇提升提供科学依据。
基金项目
武汉工程大学研究生教育创新基金项目(CX2022550),武汉工程大学高层次人才启动项目(21QD02)。
参考文献
NOTES
*通讯作者。