摘要: 通过2019年11月(秋季)和2020年5月(春季)海阳市海域两个航次的底栖生物调查,共鉴定出底栖生物62种,其中多毛类44种(71.0%),甲壳类12种(19.4%),棘皮动物2种(0.03%),软体动物3种(0.05%),纽形动物1种(0.02%)。秋季发现2种优势种,中蚓虫(Mediomastus californiensis)为第一优势种;春季发现5种优势种,独指虫(Aricidea fragilis)为第一优势种。2019年秋季底栖生物丰度平均值为432.22 ind./m
2,显著低于2020年春季1271.37 ind./m
2 (p < 0.01)。2019年秋季底栖生物多样性和丰富度指数在近岸海域较低,均匀度指数则在近岸海域及千里岩岛邻近海域较高;2020年春季底栖生物多样性和丰富度指数在近岸海域较低,均匀度指数则在近岸海域较高。春季的多样性和丰富度指数显著高于秋季(p < 0.05),但两季均匀度指数无显著差异(p > 0.05)。
Abstract:
Two cruise survey samplings of benthic organisms were conducted in the marine area of Haiyang in November 2019 and May 2020, corresponding to autumn and spring. We identified 62 species and 44 of them belong to Polychaeta, accounting for 71.0%; 12 species belong to Crustacea, accounting for 19.4%; 2 species belong to Echinodermata, accounting for 0.03%; 3 species belong to Granulifusus kiranus, accounting for 0.05%; 1 species belongs to Nemertina, accounting for 0.02%. Two species were dominant species in autumn and Mediomastus californiensis was the first dominant species. Five species were dominant species in spring and Aricidea fragilis was the first dominant species. The average abundance value of benthic organisms was 432.22 ind./m2, significantly lower than the value of 1271.37 ind./m2 in spring (p < 0.01). The nutrients, which affected the growth of benthic diatom and thus affected the abundance of benthic organisms, were higher in spring, because the seasonal changes may be the reason of the difference between them. In autumn 2019, Shan-non-Wiener diversity and Margalef’s richness indices of benthic organisms were lower in the coastal sea, while the Pielou evenness index was higher in the coastal waters and the waters near the Qi-anliyan islets. In the spring of 2020, the diversity and richness indices of benthic organisms were lower in the coastal waters, while the evenness index was higher in the coastal waters. The diversity and richness indices in spring were significantly higher than those in autumn (p < 0.05), but the difference of evenness index between the two seasons was not significant (p > 0.05).
1. 引言
底栖生物是海洋生态系统的重要组成部分,对海洋生态系统的物质循环和能量传递有重要作用,通过生物沉降和扰动作用影响海洋环境的修复过程 [1] 。底栖生物活动能力弱、地域性强,其种类组成和群落结构对当地环境状况有指示作用 [2] [3] 。因此开展底栖生物调查监测,并分析其群落结构及时空演化,对维系海洋生态系统的稳定健康及服务功能具有重要意义 [4] 。
海阳市所辖海域东起与乳山市交界的琵琶岛官厅咀,西至与莱阳市交界的丁字湾,是全省首批海洋生态文明示范区,海域内包括千里岩、土埠岛等7个海岛,以及丁字湾、马河港等8处港湾。海域拥有乳山–海阳沿岸等渔场,自然条件好、渔业资源丰富 [5] 。目前关于海阳市海域底栖生物的研究鲜有报道,因此开展这方面的研究可对海阳市海域的生态保护和渔业健康发展提供基础数据支持。
本研究基于2019年11月(秋季)和2020年5月(春季)在海阳市海域调查的底栖生物数据,分析底栖生物种类组成、丰度和生物量分布以及群落结构的时空变化,为保护该海域生物多样性和海洋生态系统服务价值评价提供基础资料。
2. 材料和方法
2.1. 研究区域和数据采集
研究区域位于海阳市所辖海域(图1蓝色部分)。调查时间为2019年11月(秋季)和2020年(春季)。其中2019年11月(秋季)设置23个调查站位,2020年(春季)设置25个调查站位。底栖生物采用抓斗式采泥器获取,每个站位采样4次以上。样品经5%福尔马林固定保存。带回实验室后,进行种类鉴定与计数,底栖生物密度标准化为个/m2(ind./m2)和g/m2作为调查水域的现存量指标。样品的采集和分析均按照《海洋调查规范》(GB/T12763-2007) [6] 进行。

Figure 1. Spatial distribution of benthic species at benthic survey stations
图1. 底栖生物调查站位的种类数空间分布
2.2. 分析方法
采用香农–威纳(Shannon-Weaner)多样性指数(H′) [7] 、Margalef物种丰富度指数(D) [8] 、Pielou均匀度指数(J′) [9] 及各物种的优势度(Y)作为评价底栖生物群落的指标,计算公式如下:
(1)
(2)
(3)
式中:S为样品中的总种数,Pi为第i种个体数与样品总个体数比值,N为样品中的生物个体数,Hmax为log2S,表示多样性指数的最大值。
(4)
式中:ni为群落中即所有调查站位第i种的个体总数,N为群落中所有物种的总个体数,fi为第i种个体在各样品中的出现频率。在调查海域以优势度Y > 0.02的种类定为优势种 [10] 。
采用K-W秩和检验方法检验不同季节底栖生物的种类数、丰度、生物量以及群落结构参数(H′、D′、J′)差异的显著性。数据分析在R软件平台上完成,群落结构指数使用Vegan包计算。
3. 结果
3.1. 底栖生物种类组成
海阳市海域底栖生物资源调查共鉴定出62种,隶属5门,其中多毛类占71.0%,甲壳类占19.4%,棘皮动物占0.03%,软体动物占0.05%,纽形动物占0.02%。2019年秋季鉴定底栖生物23种,2020年春季鉴定底栖生物50种。各季节底栖生物组成如图2所示。
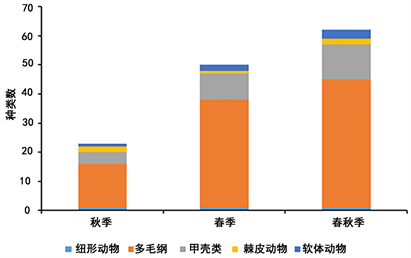
Figure 2. Number of benthic species in different seasons
图2. 不同季节底栖生物种类数
3.2. 底栖生物群落优势种
通过计算种类优势度(Y),以调查海域优势度(Y) > 0.02的种类定为优势种。2019年秋季优势种为中蚓虫和索沙蚕,中蚓虫为第一优势种,优势度为0.110 (表1),2020年春季优势种为独指虫、花冈钩毛虫、昆士兰稚齿虫、寡鳃齿吻沙蚕、双唇索沙蚕,独指虫为第一优势种,优势度为0.098 (表2)。

Table 1. The dominant species and dominance of benthic organisms investigated in autumn of 2019
表1. 2019年秋季调查海域底栖生物优势种和优势度
Table 2. The dominant species and dominance of benthic organisms investigated in spring of 2020
表2. 2020年春调查海域底栖生物优势种和优势度
3.3. 底栖生物丰度和生物量分布
2019年秋季调查海域底栖生物的丰度平均值为432 ind./m2。其中H19站位丰度最高,为3176 ind./m2,最低值157 ind./m2位于H20站位(图3(a))。调查海域底栖生物生物量平均值为28.88 g/m2,其中H19站位有最高值644.35 g/m2,H20站位有最低值0.35 g/m2 (图4(a))。2020年春季调查海域底栖生物的丰度平均值为1271 ind./m2。其中H21站位有最高值4314 ind./m2,最低值294 ind./m2位于H15 (图3(b))。底栖生物生物量平均值为194.73 g/m2,H21站位有最高值4454.71 g/m2,最低值1.57 g/m2位于H12站位(图4(b))。2019年秋季和2020年春季的底栖生物丰度和生物量空间分布图均显示近岸海域底栖生物密度较低,千里岩岛附近的外部海域底栖生物密度较高。且2019年秋季底栖生物丰度显著低于2020年春季(p < 0.01);两季生物量无显著差异(p > 0.05)。
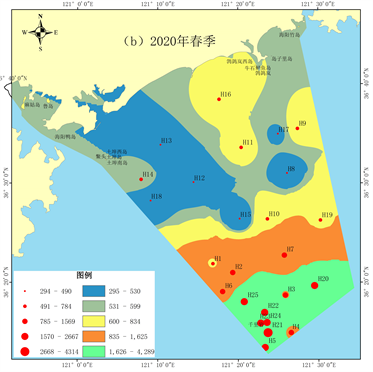
Figure 3. Spatial distribution of benthic abundance (ind./m2)
图3. 底栖生物丰度空间分布(ind./m2)

Figure 4. Spatial distribution of benthic biomass (g/m2)
图4. 底栖生物生物量空间分布(g/m2)
3.4. 底栖生物群落结构
2019年秋季海阳市海域底栖生物多样性指数平均值为1.21。多样性指数最高值出现在H2站位,最低值出现在H19站位;近岸海域底栖生物多样性指数较低(图5(a))。2020年春季底栖生物多样性指数平均值为1.74,显著高于秋季(p < 0.05)。最高值出现在H22站位,最低值出现在H8站位。近岸海域底栖生物多样性指数较低,千里岩岛附近海域的底栖生物多样性指数较高(图5(b))。
2019年秋季均匀度指数平均值为0.92,H19站位均匀度指数最低,H3、H6、H12、H15、H16、H21、H23这7个站位的均匀度指数为1.0。近岸海域及千里岩岛邻近海域底栖生物的均匀度指数较高(图6(a))。2020年春季均匀度指数平均值为0.93,与秋季均匀度指数无显著差异(p > 0.05),H23站位均匀度指数最低,H9、H15号站位的均匀度指数为1.0。近岸海域底栖生物的均匀度指数较高,千里岩岛邻近站位的底栖生物均匀度指数较低(图6(b))。
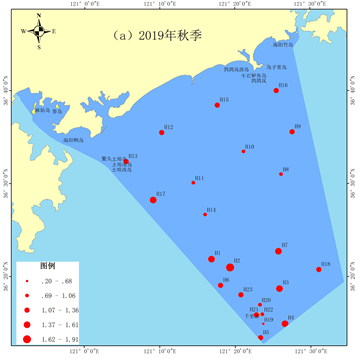
Figure 5. Spatial distribution of diversity index
图5. 多样性指数空间分布
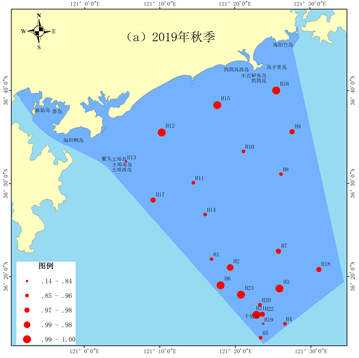
Figure 6. Spatial distribution of evenness index
图6. 均匀度指数空间分布
2019年秋季丰富度指数平均值为0.39。最高值出现在H2站位,最低值出现在H10、H14和H22站位。近岸海域及千里岩岛邻近海域底栖生物的丰富度指数较低(图7(a))。2020年春季丰富度指数平均值为0.87,显著高于秋季(p < 0.05)。最高值出现在H22站位,最低值出现在H8站位。近岸海域底栖生物的丰富度指数较低,千里岩岛邻近海域底栖生物丰富度指数较高(图7(b))。
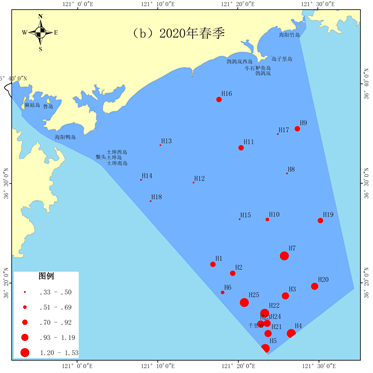
Figure 7. Spatial distribution of richness index
图7. 丰富度指数空间分布
3.5. 聚类分析结果
2019年秋季聚类分析结果显示各调查站位底栖生物群落结构可分为5类,总体趋势为近岸调查站位归并为一大类,千里岩岛邻近海域调查站位归并为一大类。H19站位单独为一类,H1、H15、H17站位分为一类,H3、H21分为一类(图8(a))。距离较近调查站位底栖生物群落结构相似度较高。2020年春季聚类分析结果显示底栖生物群落结构可分为4类。千里岩岛邻近海域调查站位划分为一类;调查区域东北侧H16、H17、H19站位分为一类,西南侧H12、H14、H18站位划分为一类;剩余调查站位划分为一类(图8(b))。
4. 讨论
4.1. 底栖生物空间分布变化
研究结果表明,千里岩岛临近的外部海域底栖生物丰度、生物量密度、生物多样性指数和丰富度指数均高于近岸海域。张亮等 [4] 以及蔡丽萍等 [11] 分别对千里岩岛海域以及舟山近海底栖生物结构进行了调查,指出底栖生物的密度、生物量与一些沉积物中的污染因子,如铜、镉等重金属元素呈负相关,而重金属主要在近海沉积,从近海向外,重金属含量依次递减,呈舌状分布 [12] ,这些因素可能是导致外海底栖生物丰度高于近岸海域的主要原因。此外,千里岩岛海洋生态系统省级自然保护区的建立,对千里岩岛海域进行了有效保护,降低了海域污染水平,减少人为因素对海洋生态系统的影响 [13] [14] ,因此千里岩岛临近海域底栖生物的丰度和多样性得到了提升,群落结构更加稳定。本文在前人研究的基础上进一步分析了造成底栖生物分布时空差异以及外海群落结构稳定性的深层次原因,突显了自然保护区在保护生物多样性方面的作用。
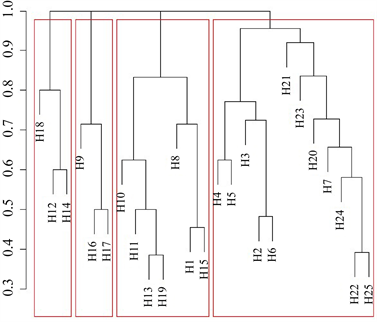
Figure 8. Cluster analysis of benthic organism
图8. 底栖生物聚类分析
4.2. 底栖生物季节分布变化
春季底栖生物的种类数量、丰度、生物量及丰富度指数显著高于秋季,这是由于季节变化导致的海洋环境变化引起的。千里岩岛近海大气湿沉降的营养盐浓度有明显的季节性,春冬两季浓度较高,夏秋两季较低 [15] 。而春季较高浓度的营养盐有利于底栖藻类的生长,提高了海域的初级生产力,使得底栖生物的食物来源更加充分,进而提升了底栖生物在春季的丰度。张亮等 [4] 分析了2015年秋季千里岩岛水产种质资源保护区海域大型底栖生物群落结构特征,本研究扩大了研究的时间和空间尺度,系统全面的分析了环境变化对大型底栖生物群落结构的驱动作用,研究成果为海阳市海域的开发利用和管理保护提供基础数据支持。
4.3. 底栖生物种类组成变化
底栖生物的种类组成变化与海洋生态系统稳定性密切相关。研究结果表明季节性的环境变化对底栖生物的多样性和优势种有显著影响。2019年秋季,中蚓虫和索沙蚕成为优势种,而在2020年春季,独指虫等多种优势种出现。这种特定优势种的变化可能与其生理特征和环境适应性有关。中蚓虫生活史特点为高繁殖力和适应低氧环境,这使其在秋季低氧的海域中占据优势。而独指虫具有较高的生长速率和适应富营养盐的能力,因此在春季的高营养盐环境中获得了竞争优势。多样的底栖生物群落通常有助于维持生态系统的稳定性 [16] 。同时,种类组成的变化也可能影响生态系统的抗干扰能力和竞争关系 [17] 。因此,深入研究底栖生物群落结构的季节性变化对了解底栖生物群落对环境变化的响应,以及制定该海域生态保护策略具有重要意义。
5. 结论
本研究基于2019秋季和2020春季两个季度的海阳市海域底栖生物调查数据,分析了底栖生物群落结构特征及与环境因子的关系,主要结论为:
1) 海阳市海域底栖生物群落结构和时空分布研究结果表明:与近岸海域相比,千里岩岛附近海域底栖生物的多样性、均匀度和丰富度指数均较高,且底栖生物群落结构更加稳定。这一结论印证了千里岩岛海洋生态系统省级自然保护区在保护该海域生物多样性方面的重要作用。
2) 底栖生物群落结构和优势种的季节性更替变化结果表明:环境因素对底栖生物群落结构的驱动作用,进一步说明底栖生物对环境变化的指示作用。因此,开展海阳市海域底栖生物长期监测,掌握底栖生物群落变化规律,对制定海域生态管理和保护策略具有重要指导作用。