1. 前言
植物多样性,是指地球上的植物及其与其他生物、环境所形成的所有形式、层次、组合的多样化,不仅与整个自然界的生物多样性相关,也与人类的生产、生活有着直接或间接的联系,直接影响人们的生活质量。
建立自然保护区是植物多样性保护的有效途径。广西九万山自然保护区始建于1982年,于2007年获准晋升为国家级自然保护区,是以保护涵养水源的天然常绿阔叶林及珍稀濒危野生动植物栖息地为主要目的的森林生态系统保护区。保护区的建立使得繁茂的森林和丰富的物种资源得到了较好的保护与发展。1989年至1990年,中国科学院植物研究所与柳州地区林业局进行了一次较大规模的植物资源考察,发现该区域有维管植物3000种以上,出版了《广西九万山植物资源考察报告》;1989~1990年,贾渝等研究了广西九万山藓类植物区系,探讨其对划分热带、亚热带分界线的意义;2001年至2002年,广西植物研究所对融水境内的九万山保护区进行了考察,结果发现其生物多样性丰富度较高;2003年,广西林业厅保护站、广西植物研究所联合多个单位组成考察队,全面系统地考察了九万山自然保护区,再次证明了九万山生物多样性及其他资源的丰富性,编著了《生物多样性关键地区——广西九万山自然保护区科学考察集》。多次的考察结果均表明,九万山自然保护区是一座不可多得的天然种质资源宝库,具有特殊的保护价值。在已有的调查研究中,关于保护区内固定样地植物群落结构的数量特征及其植物演替的相关研究较少。本项目基于2011年7月至8月野外实地调查的数据,为研究样地植物群落动态变化监测提供基础材料和数据,通过统计群落的个体数量指标和综合数量指标,分析不同群落的物种组成结构及其群落多样性与环境因子的相互关系,并与国内相同气候区的自然保护区的植物多样性调查数据进行对比分析,旨在进一步了解九万山自然保护区植物多样性的分布特征,以利于植物资源的有效保护和管理,为生态系统服务功能的发挥和区域经济发展做贡献。
2. 材料与方法
2.1. 自然概况
九万山国家级自然保护区位于广西壮族自治区北部,地跨融水苗族自治县、环江毛南族自治县及罗城仫佬族自治县,属于长江和珠江水系分水岭山脉的南缘部分,东经108˚35′32″~108˚48′49″,北纬25˚01′55″~25˚19′54″,总面积25,212.8 hm2。该保护区处在中亚热带季风气候区内,其西北高东南低的特殊地势为植被提供了良好的水热条件,年平均日照时数为1000~1200 h,年平均气温12.0℃~17.1℃,年降水量1600~2100 mm,年蒸发量1384.7~1466.0 mm。地貌类型属于侵蚀褶皱深–中等切割中山,中山地貌特征显著,北面山高谷深,地形崎岖,最高海拔1693 m,坡度多为50˚上下;南面山势稍为缓和,坡度一般为30˚~45˚,复杂的地貌很大程度影响着植被的生长分布。土壤类型有山地红壤、山地红黄壤、山地黄壤、山地黄棕壤和山地矮林土五大类。保护区内以中亚热带典型常绿阔叶林面积最大、分布最广,较高海拔地带有中山常绿落叶阔叶混交林和中山针阔混交林,山顶和山脊地段出现矮林。据报道,九万山国家级自然保护区已知维管植物有2754种,隶属236科980属。其中蕨类植物有48科107属314种,裸子植物有7科12属19种,被子植物有181科861属2421种,国家重点保护植物有22种 [1]。
2.2. 调查方法
在九万山国家级自然保护区管理局相关技术人员的帮助下,根据调查研究的目的、保护区重要监测(保护)物种的分布、森林植物群落的主要类型、功能区的划分、自然环境的影响及人为干扰的程度,经过反复的实地考察和技术论证,筛选并确定了24个20 m × 50 m的永久监测样地(其中核心区10个,缓冲区和试验区各7个),样地设置情况见图1和图2。调查时把每个大样方又分为10个10 m × 10 m的乔木小样方,个别地段地形复杂不宜设样地。利用地质罗盘仪、经纬仪及全球卫星定位仪(GPS)对每个样地的四个边角和中心点进行了定位并测定了坐标参数(经纬度及海拔高度),测定了每个监测样地的坡度、坡向、植被覆盖率、郁闭度、土层厚度等基本参数。根据植物生态学野外调查方法,对每个监测样地植物群落的物种多样性指数进行调查,对样方内所有胸高直径(DBH) ≥ 5 cm的乔木进行每木检尺,分别记录其编号、种名、胸径(DBH)、树高、冠幅等,并挂金属(铝合金)牌做永久标志。灌木或草本植物的多样性调查则按照梅花形布点方法,在大样方中划定5个大小为5 m × 5 m或1 m × 1 m的小样方,分别记录其编号、种名、盖度、高度和株数等,对灌木挂铝合金牌做永久标志。
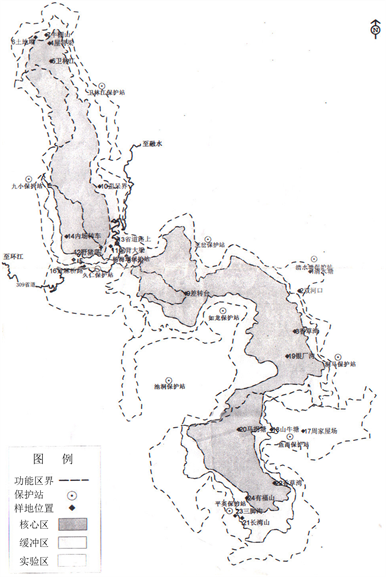
Figure 1. The location of the Jiuwan Mountain National Nature Reserve in Guangxi and the survey plots
图1. 广西九万山国家级自然保护区及调查样地位置图示
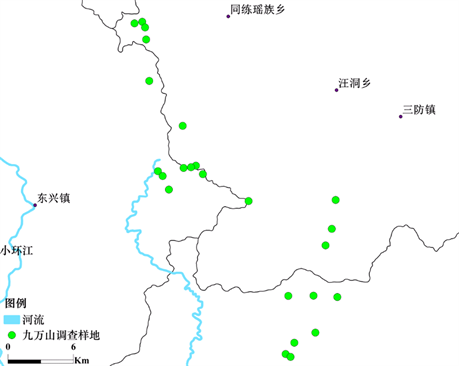
Figure 2. Distribution of plots for biodiversity survey in Guangxi Jiuwan Mountain Nature Reserve
图2. 广西九万山自然保护区生物多样性调查样地分布
2.3. 数据处理方法
2.3.1. 群落层片水平上的多样性量度
公式如下:
(乔木层)重要值
(灌木层、草本层)重要值
Shannon-Wiener指数
Simpson指数
均匀度PieLou指数
其中,S为丰富度,即种的数目;Ai为第i种物种的相对重要值,即同一层片中第i种植物的重要值除以所有植物的重要值之和。
2.3.2. 群落总体水平多样性量度
本文的群落多样性是所有乔、灌、草各物种一起参与多样性的计算。采用给定加权参数法确定物种的相对重要值Pi:
其中,Ai1、Ai2、Ai3分别为物种在各层片中的相对重要值,Pi1、Pi2、Pi3为给定加权参数后物种在各层片中的相对重要值,W1、W2、W3分别为给定的乔木层、灌木层、草本层物种权重系数,结合各片层在群落中的生态作用大小采用0.5、0.25、0.25 [2]。若某一物种仅在乔木层出现,则其在群落中的重要值为Pi1,若在的乔木层和灌木层均有出现,则其在群落中的相对重要值为Pi1 + Pi2。群落α多样性指数计算公式同2.3.1,其中Ai改为Pi。
2.3.3. 样地归类分组
坡向划分以北为起点,顺时针方向旋转,按照阴坡(315˚~45˚),阳坡(135˚~225˚),半阴坡(45˚~90˚和270˚~315˚),半阳坡(90˚~135˚和225˚~270˚),将24个样地中具有典型方位的21个样地分为半阳坡、半阴坡、阴坡三组,每组均7个样地。样地坡度分3个等级,40˚~60˚为险坡,30˚~40˚为陡坡,0˚~30˚为缓坡,同样将24个样地中具有典型坡度的21个样地分三组,每组均7个样地。
2.3.4. 统计分析
利用Excel 2003或Origin 8.0进行数据处理或曲线拟合,运用单因素方差分析比较均值差异。根据小概率原理,取α = 0.05,α = 0.01为显著水平和极显著水平,P < 0.05为显著,P < 0.01为极显著。
3. 结果与分析
3.1. 植被群落结构特征
3.1.1. 样地基本情况
本研究的24个野外调查样地基本情况见表1。

Table 1. The natural profile of the surveyed plots
表1. 调查样地的自然概况
3.1.2. 群落结构特征
保护区样地森林群落由于受到人为干扰程度小,植被十分茂密,原生性较强,物种组成以壳斗科Fagaceae、樟科Lauraceae、山茶科Theaceae、杜鹃花科Eticaceae、冬青科Aquifoliaceae等物种居多。群落成层结构明显,分为乔木层、灌木层、草本层3层,乔木层优势显著。乔木层林木高度多为10~15 m,部分个体高度可达25 m以上,胸径在5~116 cm之间,平均胸径7.98~15.15 cm,依据林木高度小于10米划为亚乔木层,第1乔木层盖度变化范围10%~80%,亚乔木层盖度变化范围20%~70%。样地范围(20 m × 50 m)内有乔木12~42种,83~383株,平均34种,201.50株。对乔木层树种重要值进行分析,重要值较大的树种是贵州桤叶树(Clethra kaipoensis)、海南杜鹃(Rhododendron hainanense)、东南野桐(Mallotus lianus)、星毛鸭脚木(Schfflera minutistellata)等,重要值分别为37.70、34.95、25.62、24.55 (见表2,表3)。进一步分析重要值的离散性发现,各物种重要值比较接近,变异系数0.78~2.14,群落的乔木层第一优势种很少能占据绝对优势,只有在7号和9号样地中的海南杜鹃和贵州桤叶树占据较大的优势。因此,九万山原生性森林优势种不明显。
Table 2. Descriptive statistics of important values of tree layer in each plot
表2. 各样地乔木层重要值的描述统计
灌木层植株高0.5~6 m不等,多为2~5 m,个别乔木幼树(DBH < 5 cm)达到10 m,该层盖度变化范围15%~80%。物种组成多由摆竹(Indosasa shibataeoides)、匙萼柏拉木Blastus cavaleriei、杜茎山(Maesa japonica)、柃木属(Eurya)以及乔木的幼苗幼树等构成,尤以乔木的幼苗幼树居多,但其往往不占据优势。在调查范围(5 m × 5 m × 5)内有灌木18~42种,36~2019株,平均29.33种,378.33株。该层与乔木层物种数量比值在0.47~1.57之间,平均比值为0.92,灌木物种组成及盖度与乔木层有一定的关联性。对灌木层物种重要值的离散性进行分析,变异系数在0.72~1.96之间,各物种重要值十分接近,仅少数物种重要值较大,优势相对明显,如摆竹和赤杨叶(Alniphyllum fortunei)。
草本层一般高度小于1 m,物种组成及盖度受上层影响较大,物种有锦香草(Phyllagathis cavaleriei)、狗脊蕨(Woodwardia japonica)、赤车(Pellionia radicans)、光里白(Diplopterygium laevissimum)、沿阶草(Ophiopogon bodinieri)等等,蕨类植物种类较多。盖度0.5%~90%,在乔木层或灌木层盖度较大的样方中,草本层盖度多小于5%。在样格范围(1 m × 1 m × 5)内,有物种2~23种,13~353株,平均10.75种,98.75株。对草本层物种重要值的离散性进行分析,变异系数在0.60~2.11之间,各物种重要值十分接近,少数种优势明显,如锦香草、光里白。
Table 3. Dominant species in different layers of plant communities in the survey plots
表3. 调查样地植物群落不同层片的优势种
3.2. 群落多样性特征
3.2.1. 群落α多样性
在调查样地中,群落丰富度(S)范围31~80,Shannon-Wiener指数(H)范围2.8861~3.8108,Simpson指数(D)范围0.9248~0.9673,均匀度PieLou指数范围0.7947~0.8980。群落分层明显,各层α多样性指数见图3~6。
从图3~6可以看出,在调查样地中,乔木层和灌木层的丰富度数值较接近,总体上变化趋势相似,草本层丰富度最低。Shannon-Wiener指数规律性不明显,总体而言,乔木层和灌木层高于草本层。总体来说,Simpson指数变化很小,基本持平,与乔木层和灌木层相比,草本层Simpson指数变化则非常大。各样地总体及各层片的均匀度PieLou指数变化均较大。

Figure 3. Comparison of species richness of surveyed plots
图3. 调查样地物种丰富度的比较
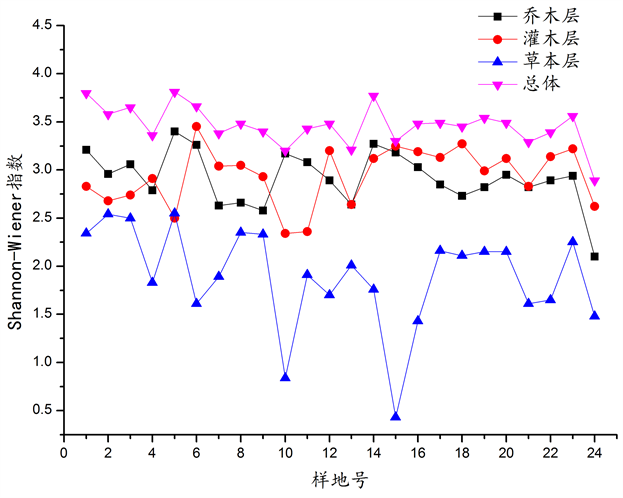
Figure 4. Comparison of Shannon-Wiener index of surveyed plots
图4. 调查样地Shannon-Wiener指数的比较
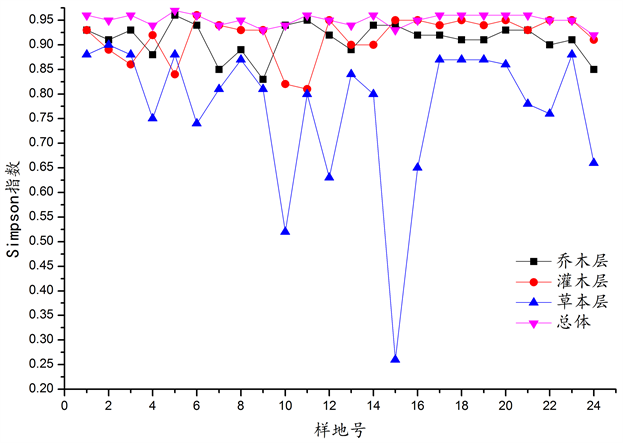
Figure 5. Comparison of Simpson index of surveyed plots
图5. 调查样地Simpson指数的比较

Figure 6. Comparison of PieLou index of evenness of survey plots
图6. 调查样地均匀度PieLou指数的比较
对24个样地各层片的α多样性指数进行均值差异分析,结果表明:各层片的丰富度的均值差异极显著,
,表现为乔木层(S = 34) > 灌木层(S = 29) > 草本层(S = 11)。
各层片的Shannon-Wiener指数和Simpson指数的均值差异都达到极显著水平,
,表现为灌木层(H = 2.941; E = 0.918) > 乔木层(H = 2.912; E = 0.912) > 草本层(H = 1.900; E = 0.773)。
各层片的均匀度PieLou指数均值差异不显著。
3.2.2. 实验区、缓冲区、核心区的多样性比较
核心区4号、13号、24号样地情况较为特别,群落特征比较特异,13号样地有明显人为干扰痕迹,多样性指数(H = 3.210)和乔木胸高断面积总和(值为1.7624 m2)均较低。4号和24号样地无明显人为干扰迹象,木材蓄积高,名列前2位。4号样地有乔木41种,383株,胸高断面积总和3.8027平方米,群落出现藤本绞杀现象,种间竞争比较激烈;24号样地有乔木12种,137株,胸高断面积总和3.4089平方米,有较大的倒木,已枯死,林密度较小。二者的显著差异在一定程度上反映核心区存在不同的群落演替阶段。
基于4号、13号、24号样地的特殊性,不考虑加入核心组。将剩下的21个样地按实际地域划分均分为核心组、缓冲组、实验组三组,并进行α多样性指数均值比较,结果表明:Shannon-Wiener指数差异达到显著水平(P = 0.037),核心组(3.583) > 实验组(3.553) > 缓冲组(3.382);Simpson指数差异达到显著水平(P = 0.011),核心组(0.960) > 实验组(0.954) > 缓冲组(0.944);丰富度和均匀度PieLou指数没有显著差异。总体来看,由于受到人为干扰少,核心组多样性程度相对较高,优势种较为突出;而受到一定人为干扰的实验组多样性也比较高,符合适度干扰可提高物种多样性的中等干扰学说。
3.2.3. 群落多样性与海拔的相关性
九万山国家级自然保护区地形相对高差比较悬殊,随着海拔的变化,乔木层优势种发生明显改变。在低海拔样地以第一优势出现多次的种有:星毛鸭脚木和米槠,在较高海拔以第一优势出现多次的种有:海南杜鹃和贵州桤叶树。仅在最高海拔的11号样地出现小叶罗汉松(Podocarpus wangii),重要值为3.33,优势不明显。
在本次调查设置的样地中,群落总体的丰富度(S)、Shannon-Wiener指数(H)、Simpson指数(D)与海拔变化并无显著关系(见图7),乔木层丰富度指数(S)、Shannon-Wiener指数(H)、Simpson指数(D)和均匀度PieLou指数(E)与海拔无明显关系(见图8)。但草本和灌木的均匀度与海拔变化存在着显著关系,在一定范围内,随着海拔的升高,草本、灌木以及群落总体的均匀度呈下降趋势,具体情况见图9,草本层R2 = 0.210,P = 0.032;图10,灌木层R2 = 0.310,P = 0.008;图11,总体R2 = 0.231,P = 0.024。

Figure 7. Changes in the overall diversity index of plant communities in the survey plots with altitude
图7. 调查样地植物群落总体多样性指数随海拔的变化
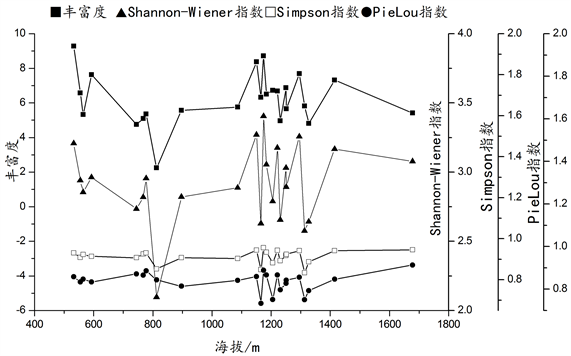
Figure 8. Variations of the arbor tree diversity index of the survey plots with altitude
图8. 调查样地乔木多样性指数随海拔的变化

Figure 9. The relationship between herb evenness and altitude in the survey plots
图9. 调查样地草本均匀度与海拔的关系
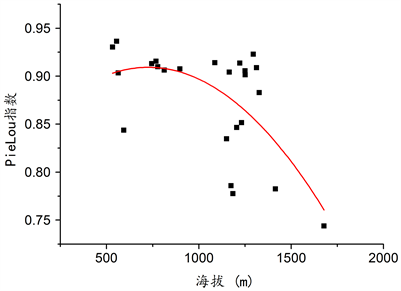
Figure 10. The relationship between the evenness of shrubs on the survey plot and altitude
图10. 调查样地灌木均匀度与海拔的关系
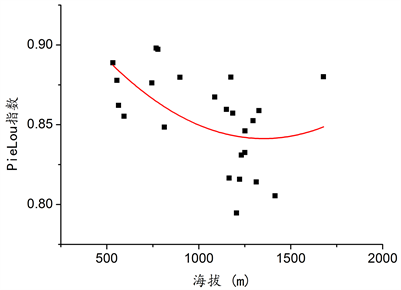
Figure 11. The relationship between the overall evenness of the community in the survey sample plot and altitude
图11. 调查样地群落总体均匀度与海拔的关系
3.2.4. 植物多样性与坡向、坡度的相关性
进行半阳坡、半阴坡和阴坡植物多样性比较发现:乔木层、草本层以及群落总体多样性与坡向无显著相关性;灌木层丰富度指数(S)与坡向相关性较为显著(P = 0.037),表现为半阳坡 > 半阴坡 > 阴坡;灌木层Shannon-Wiener指数(H)与坡向相关性极显著(P = 0.002),表现为半阳坡 > 半阴坡 > 阴坡;灌木层Simpson指数(D)、均匀度PieLou指数(E)与坡向相关性显著(P = 0.001),均表现为半阴坡 > 半阳坡 >阴坡,见图12。
进行缓坡、陡坡和险坡植物多样性比较发现:乔木层、草本层和群落总体多样性指数以及灌木均匀度PieLou指数(E)与坡度的相关性不显著;灌木的丰富度指数(S)、Shannon-Wiener指数(H)、Simpson指数(D)与坡度的相关性显著(PM = 0.002; PH = 0.008; PD = 0.041),均匀度PieLou指数与坡度相关性不明显(PE = 0.278),均表现为陡坡 > 险坡 > 缓坡,见图13。

Figure 12. Comparison of shrub diversity in plots with different slope directions
图12. 不同坡向样地的灌木多样性的比较

Figure 13. Comparison of shrub diversity in plots with different slopes
图13. 不同坡度样地的灌木多样性的比较
4. 讨论
4.1. 植物多样性情况
根据乔木层、灌木层、草本层对群落的贡献不同采用给定加权参数法,得到的总体α多样性指数与无加权参数的总体α多样性指数两组数据无显著差异(
),但使用加权参数法得到的总体α多样性指数范围较大,相比幅度变化±0.06,更容易反映样地群落多样性之间的功能差异,并强化了对比。
调查结果表明,广西九万山国家级自然保护区具有较高的物种多样性,特别是乔木的物种多样性;相比同属中亚热带气候区的国内其它几个自然保护区 [3] [4] [5] [6] (见表4),九万山国家级自然保护区植物(物种)多样性的最大特点是不同地域间的多样性变化幅度小,在一定程度上反映了九万山自然环境的优越性和良好的生态环境;在海拔高度与植物分布的关系上,表现为不同海拔出现不同的优势种,植物群落的组成和结构也有一定的差异;由于样地间存在复杂地形条件的综合影响和人为干扰等各种因素,削弱了群落的多样性指数随海拔高度变化的规律性。相对来说,海拔1100~1300 m之间的植物多样性比较高。
Table 4. Comparison of woody plant diversity in national nature reserves in the mid-subtropical climate zone
表4. 中亚热带气候区的国家级自然保护区木本植物多样性比较
备注:采用参考文献 [2] [3] [4] [5] 的相关数据,由于广东南岭Simpson指数计算公式不同,不选用;贵州茂兰和江西武夷山分别缺均匀度指数和Simpson指数,数据为空。
4.2. 环境因子与植物多样性
植物多样性与众多因素相关,其中环境因子对植物(物种)多样性有着非常重要的影响 [7] - [17]。随着海拔的升高,植物自然生长环境变得更加恶劣,比如土层厚度变薄且分布不均,土壤贫瘠,养分缺乏,风害、寒害比较严重,等等。从图7~11的多样性指数变化来看,乔木的物种多样性随海拔升高无明显变化(具体原因有待进一步研究),灌木、草本均匀度PieLou指数随海拔升高呈下降趋势 [18] [19]。不同环境因子对不同生长型植物的影响作用是有差异的,也就是说乔木、灌木、草本的主要环境限制因子不同 [20] [21] [22] [23] [24]。由于多种生态因子的综合影响,海拔对乔木多样性的影响作用在一定程度被削弱,或者由于本调查区域的海拔高度相对来说不是太高,不同样地的海拔高度差异较小,不足以使乔木的物种多样性呈现明显的规律性变化。另外,乔木有着较强的生活力,在群落占据绝对的优势地位,适应海拔等环境因子变化的能力较强,因而随着海拔升高生长状况基本不变,物种多样性变化不明显。海拔高度变化对灌木、草本的影响表现明显,其与草本、灌木均匀度的负相关关系分别达到显著和极显著水平,这可能与土层厚度和养分条件变化有较大关系 [25] [26]。以上结果,与张玲等人对黑龙江胜山保护区调查的结果,即乔木多样性与海拔呈正相关,灌木、草本与海拔无明显关系不一致 [27]。
坡向影响群落的光和水热条件,对灌木的影响很明显 [28],图12至图13中半阳坡的灌木丰富度指数、Shannon-Wiener指数明显比阴坡高,物种丰富,半阴坡介于二者中间;在分布均匀程度方面,阴坡的灌木Simpson指数和均匀度PieLou指数最低,也就是阴坡灌木分布最不均匀,优势种明显。因为不同的植物适应于一定的坡向,阴坡光和水热条件较差,对灌木生长有一定的限制,多样性低,利于耐阴性的物种形成优势。半阳坡和半阴坡接受的光照和热量相对阴坡多,有利于植物生长,因此多样性高,群落竞争激烈因而优势种不明显。
坡度通过影响土壤的厚度和理化性质等间接影响植物的生长,对灌木多样性的影响比较明显。大坡度,土层薄,营养物质易被淋失,水分少,有机质积累少,小坡度,土层较厚,水分多,有机质易于积累,理论上小坡度适于植物生长,灌木多样性较高。图13陡坡与险坡比较,陡坡坡度相对较小,灌木多样性指数相对较高,与理论相符。缓坡坡度最小,而灌木多样性指数却最低,由于坡度大小与人为干扰强弱是负相关的,缓坡人为干扰大,存在偷砍盗挖现象几率大,植被易受到破坏,以致灌木多样性指数降低 [29]。虽然陡坡和险坡坡度大,植被自然生境较差,但受到较少人为干扰,多样性仍然是比较高的。
不同坡度样地的植物多样性差异反映人为干扰降低了植物多样性 [30],但是存在适度干扰,会使多样性提高,生态系统的适应能力、抗干扰能力会增强,生产力也可能比较高 [31]。保护区核心区人迹罕至,样地间多样性指数差异较大,多样性指数均值较高,一定程度反映了自然环境的异质性 [32] [33]。实验区与人居环境邻近,样地间多样性指数差异较小,且多样性均值与核心区很相近,较高,不排除是人为干扰的结果 [34] [35] [36]。
5. 小结
广西九万山国家级自然保护区固定样地的植物多样性较高,对保护区植物多样性分布具有代表性,此次样地调查木本植物1000多种,草本植物700多种,国家重点保护植物有10多种,具有重要的保护和研究价值。该保护区海拔对灌木和草本的影响较大,海拔越高,均匀度越小。由于海拔高度差异不大或者海拔的影响作用被其他因子削弱等原因,乔木多样性随海拔变化的规律性不明显。坡向与坡度显著影响灌木多样性,向阳坡面灌木多样性指数最高,背阴坡面灌木均匀度最小,中等坡度的灌木多样性最高,小坡度存在人为干扰,灌木多样性最低。因为保护区成立时间较晚,上世纪50年代到70年代是林木采育场地,保护区内居住村落较多,人工杉木林、楠竹林、作物区较多,森林砍伐痕迹明显,保护区成立后,保护措施有力,森林恢复较好,群落到树群、树丛明显增多,受2008年冰雪灾害的影响,保护区内枯立木、断尾木、倒立木数量较多。
保护区植物多样性(物种多样性)的调查,反映了植物物种存在和人类保护的状况。同时,调查结果显示的多样性变化,可以反映环境因子的影响及人类干扰的综合作用,研究环境因子与植物多样性的关系,可以更好地指导保护区的资源管理和生物多样性保护工作,为协调自然保护区与周边社区的关系,充分发挥森林生态系统的服务功能,促进区域经济可持续发展提供理论支持。
6. 建议
1) 有必要继续开展九万山国家级自然保护区生物多样性监测固定样地建设的后续研究
由于经费和时间的限制,本项目仅进行24个样地的植物(物种多样性)调查,由于调查时大部分植物无花无果,有些植物只能鉴定到科或属,样地的选点,虽然考虑了保护区重要保护物种的分布、森林植物群落的类型、生态系统特征、人为干扰影响,野外作业可操作性等诸多因素,具有一定的地理代表性,但是物种的代表性可能还不够。调查时,虽然记录了每个样地的原始资料和相关数据,且每个样地均埋设了水泥标桩进行标识,并给每株乔木和按梅花形设置的5个小样格的灌木挂牌标记,但十年时间已经过去,至今仍未对固定样地植物的生长和数量动态进行跟踪调查或植物群落演替研究,希望保护区和相关专家继续开展后续研究,使“24个固定监测样地”的植物生长情况、物种变化、群落演替材料更加翔实,以便更准确地预测保护区植物群落的变化,采取更有针对性的保护措施。
2) 生物多样性保护和自然保护区管理,应当考虑社区群众的利益
当下,我国正在积极推行林权制度改革,在林权所有制下放到户的今天,由于历史的原因,保护区与其周边的居民可能存在一些山界纠纷。由于经济利益的驱使,有的群众认为,自然保护区的成立,使他们的部分山头被占了,经济利益受到一定的损害。不少居民只顾眼前,不看长远利益的事累见不鲜,珍稀树种如南方红豆杉(Taxus chinensis)、珍珠罗汉松(Podocarpus brevifolius)、红花荷木(Rhodoleia championii)、润楠(Machilus pingii)、香楠(Aidia canthioides)、金丝楠(Phoebe sheareri)、紫檀(Pterocarpus indicus)、麻栎(Quercus acutissima)等惨遭偷盗和砍伐。事实上,自然保护区的成立和建设,并不是当地群众贫困的根本原因,但是,村民的脱贫致富和自然保护区生物多样性及生态保护是目前保护区工作的“两难问题”和迫切需要解决的问题,如何协调自然保护区与社区群众的关系,使社区群众能够支持并自觉参与自然保护区的有关工作,从而达到双赢的目的,无疑是当地政府和自然保护区管理部门必须认真思考的现实问题。在严格保护的前提下,保护区能否利用优越的自然条件和优质环境资源,通过各种渠道引进种子基金,给群众提供种源,组织开展实用技术培训,在其居住地周边进行林下种养,帮助当地群众脱贫致富?群众生活富裕了,再加上其他的惠民措施,自然保护区的资源环境保护工作一定会更好,人与自然更加和谐相处。
3) 应当根据生态建设及主体功能区的划分进行科学的管理
广西九万山国家级自然保护区是重要的水源林区,其周边农村的生活污染和农业污染也是普遍现象,因此,有必要在保护区周边的居住区建立垃圾处理池,尽量减少或杜绝使用除草剂、杀虫剂及其他化学药剂,适度控制速生桉的种植规模及种植面积,避免水源污染及造成其他危害。
基金项目
九万山国家级自然保护区生物多样性监测固定样地(一期)建设项目资助;广西高校重点实验室资助项目(桂教科研[2010]6号)。桂西北地方资源保护与利用工程中心资助项目(桂教科研[2012]9号);广西高等学校特色课程及专业一体化建设基金资助项目(桂教高教[2011]102号);广西教育科学“十二五”规划2013年度立项B类课题——服务广西区域经济发展的民族地区高校科技创新平台建设探索与实践(2013B026)。
NOTES
*通讯作者。